
БОЛЬШАЯ СОВЕТСКАЯ ЭНЦИКЛОПЕДИЯ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ
|
БОЛЬШАЯ СОВЕТСКАЯ ЭНЦИКЛОПЕДИЯ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ |
МХИ, наземные или реже пресноводные автотрофные растения, объединяемые в отдел мохообразных, включающий наиболее примитивные высшие растения. M. подразделяют на 3 класса: антоцерото-вые, печёночные и лиственные, или листостебельные. Все они сравнительно просто организованные, многоклеточные, б. ч. многолетние (реже - однолетние) растения вые. от 1 до 50 ел с частичным обособлением ассимиляционной, водо-проводящей и механич. тканей; обладают единым циклом развития, различающимся в пределах отд. групп нек-рыми особенностями. Цикл развития M. сопровождается обособлением диплоидного спорофита (видоизменённого спорангия) на гаплоидном гаметофите. В результате половое и бесполое поколения развиваются совместно на одном растении, т. е. чередование поколений у M. носит несколько условный характер. Из споры у M. вырастает ветвистая, многоклеточная нитчатая или пластинчатая протонема, способная к фотосинтезу. Вследствие развития на протонеме многочисл. почек для M. характерны групповые формы роста (дерновинки, куртинки, подушки), что помогает переносить неблагоприятные условия и способствует вегетативному размножению. У одних M. из почек вырастают пластинчатые слоевища, у других - радиально- или двусторонне-симметричные побеги. Слоевища и побеги могут быть обоеполыми или однополыми; соответственно M. бывают обоеполыми или разнополыми, однодомными и двудомными. Органы полового размножения - антеридии и архегонии - чаще располагаются группами среди многоклеточных стерильных нитей - парафиз и окружены особыми листовидными выростами. В антеридиях (продолговатых мешочках на ножке с однослойной оболочкой) образуются двужгутиковые сперматозоиды, для передвижения к-рых к яйцеклетке необходима вода. В нижней расширенной части архегония помещается яйцеклетка. Оплодотворение и дальнейшее развитие зиготы происходит в архегонии. Из зиготы за неск. месяцев развивается спорогон - орган бесполого размножения (спороноще-ния), в значит, степени утерявший самостоятельность. Зелёное стеблеобраз-ное тело молодого спорогона затем становится жёлтым, коричневым или тёмно-красным и дифференцируется на верх, спороносную часть - коробочку и нижнюю - ножку со стопой, к-рая врастает в ткань материнского растения. При образовании спор из археспория (споро-генной ткани) происходит мейоз. Зигота и спорогон у M. диплоидны и составляют бесполое поколение - спорофит. Протонема, слоевищные и олиственные гаметофоры гаплоидны и относятся к половому поколению - гаметофиту; в основном на них ложатся функции авто-трофного питания. Благодаря этому и способности гаметофита к вегетативному размножению цикл развития M. длительное время может происходить без образования спорофита (у нек-рых видов спорогоны неизвестны). Гаметофиты M. морфологически разнообразны, однослойные листовидные выросты стебля (филлоиды) отличаются большим варьированием признаков. Спорогоны же во многих группах обнаруживают значит, сходство. На стеблях олиственных M. образуются также хлорофиллоносные листо- или нитевидные выросты - пара-фил лии, булавовидные волоски, вторичная протонема и нитевидные ризоиды, к-рые, как войлоком, могут обволакивать наземные или подземные части растений. Ризоидами слоевища и стебли прикрепляются к субстрату или плотно соединяются между собой. У M. установлены апоспория и апогамия, гибридизация и полиплоидия, что даёт основания для признания их участия в видообразова-тельных процессах. Наряду с этим у M. распространено формирование клонов, что способствует сохранению местных адаптивных изменений. Антоцеротовые M. (Antho-cerotae)-своеобразная группа из 2 сем. с 6 родами, объединяющими св. 300 видов, распространённых преим. в тропиках. В СССР 4-5 видов. Слоевище у них обычно лопастное или розетковидное с погружёнными в него антеридиями и архегониями. Спорогон, как правило, вытянутый, стручкови дный, способный удлиняться благодаря меристеме, расположенной в его нижней части, с колонкой в середине и с устьицами в наружных стенках; у основания окружён обёрткой и раскрывается 2 створками сверху вниз. Кроме спор, образуются одно- или многоклеточные бесплодные короткие нити - элатеры, способствующие распространению спор. В клетках слоевища и спорогонов содержится 1 или неск. хро-матофоров с пиреноидами. Печёночные M., или печёночники (Hepaticae), подразделяются на 2 подкласса: маршанциевые и юнгер-манниевые. В отличие от других M., у печёночников протонема слабо развита, недолговечна и на ней обычно развивается одно слоевищное или листостебель-ное растение дорзо-вентральной структуры; в клетках гаметофита содержатся одно или неск. масляных телец постоянной формы; ризоиды одноклеточные. Спорогон чаще без хлоропластов, лишён колонки, и на его стенках отсутствуют устьица. У нек-рых маршанциевых обнаруживается сходство с антоцеротовыми, поэтому антоцеротовых рассматривали как подкласс печёночных M. Гаметофит печёночников отличается большим разнообразием, спорогоны однотипны. Маршанциевые печёночники (Marchantiidae) - слоевищные формы, обычно в виде дихотомически ветвящихся маленьких розеток или крупных лентовидно стелящихся слоевищ, прикрепляющихся к почве ризоидами; муж. и жен. гаметангии погружены в ткань слоевища или приподняты на особых подставках. Преим. напочвенные растения, редко скальные или водные, распространены по всему земному шару, особенно богато представлены в тропиках. Известно 16 семейств с 35 родами, объединяющими ок. 420 видов, включая ископаемую Naiadi-ta из мезозоя. Наиболее примитивны M. из порядка сферокарповых с просто устроенным слоевищем, иногда с масляными тельцами в особых клетках, с однослойной стенкой спорогона и с питат. клетками вместо элатер. Для M. порядка маршанциевых характерно сложное строение слоевища с разделением на основную и ассимиляционную ткань, с масляными тельцами в особых клетках и со скле-ренхимными волокнами (у нек-рых видов). Слоевища часто с воздушными камерами и воздушными устьицами, с нижней стороны с брюшными чешуйками и одноклеточными гладкими и язычковыми ризоидами. Спорогон обычно более сложного строения. Среди них рич-чиевые рассматриваются как вторично упрощённые, с гаметангиями и спорого-ном, развивающимся внутри мелких, чаще розетковидных слоевищ. Юнгерман-ниевые печёночники (Jungermanniidae) включают группу слоевищных и олиственных растений, в вегетативных клетках к-рых, как правило, по нескольку масляных телец; ризоиды гладкие, одноклеточные. Эпифитные, напочвенные и наскальные растения особенно богато представлены в субтропиках и тропиках. 45 сем. объединяют св. 240 родов, включающих ок. 9 тыс. видов. У представителей порядка метцгериевых слоевище одно- или многослойное, недифференцированное (у нек-рых с брюшными или спинными, иногда листовидными чешуйками и стеблями с листовидными лопастями); коробочка двух-или многослойная. Растения порядка гапломитриевых прямостоячие, с трёхрядным листорасположением и коробочками, имеющими однослойные боковые стенки. Наиболее многочисленны и разнообразны M. порядка юн-германниевых, для к-рых характерны 2 ряда цельных или лопастных боковых листьев без жилки и более мелкие (иногда отсутствуют) брюшные листья - т. н. амфи-гастрии; стенки коробочки многослойные. Лиственные M., или листо-стебельные M. (Musci), подразделяются на 3 подкласса: сфагновые, андре-евые и бриевые. Растения радиально-, редко двусторонне-симметричные, спирально олиственные; листья цельные, зубчатые или по краю реснитчатые, однослойные, реже дву- или многослойные, с жилкой или без жилки; с многоклеточными, ветвистыми ризоидами. Спорогоны верхушечные или боковые, разнообразные по форме, строению и окраске. Ок. 15 тыс. видов (по др. данным, до 25 тыс. видов), объединяемых в 660 родов. Сфагновые, или торфяные, M. имеют крупные, прямостоячие стебли с пучковидно расположенными ветвями, на верхушке собранными в головку; беловато-зелёные, жёлтые, бурые или красноватые. Листья однослойные, веточные и стеблевые, без жилки, из чередующихся хло-рофиллоносных и водоносных клеток. Спорогон состоит из шаровидной коробочки с крышечкой и недоразвитыми устьицами, без ножки, со стопой, вросшей в безлистную удлинённую архегониальную веточку стебля. Споровый мешок сводообразно покрывает верх, часть колонки. Распространены по всему земному шару; преобладают на болотах, в тундре, во влажных лесах. Андреевые M.- мелкие красно-бурые (до чёрных), с однослойными листьями; растут на скалах подушечками. Коробочка удлинённо-яйцевидная, на ложноножке, раскрывается 4-8 створками, без устьиц, со споровым мешком в виде свода над верх, частью колонки. Распространены высоко в горах, а также в сев. широтах. Бриевые M.- наиболее обширная, разнообразная группа. Коробочка обычно на настоящей ножке, с устьицами, чаще после опадения крышечки открывается наверху широким отверстием - устьем. У многих по краю устья образуется 1- 2 ряда выростов - зубцов (наз. простым или двойным перистомом), к-рые служат для рассеивания спор. Колонка, как правило, пронизывает споровый мешок. (Цикл развития см. Лиственные мхи.) M. распространены повсеместно. Поселяются повсюду, кроме морей, засоленных почв и местообитаний, скрытых под ледниками или подверженных сильной эрозии. Антоцеротовые и печёночные M. распространены в основном в странах с тропич. или умеренным влажным климатом; только немногие приспособились к произрастанию в сухих местах. Лиственные M. растут почти повсеместно, однако наибольшего развития достигают на увлажнённых местах, в лесах, тундре; на болотах M. образуют осн. массу торфяных залежей. Интенсивно развиваясь, M. способствуют заболачиванию почв, ухудшают качество лугов и др. с.-х. угодий. В связи с антибиотич. свойствами нек-рых M. их применяют иногда как перевязочный материал. Используют в качестве подстилки для скота (преим. сфагновые M.), в строит, технике для изготовления плит и др. Лит.: Флора споровых растений СССР, т. 1, 3, М.-Л., 1952-54; С а в и ч - Л ю-б и цк а я Л. И., Смирнова 3. H., Определитель сфагновых мхов СССР, Л., 1968; их же, Определитель листостебель-ных мхов СССР. Верхоплодные мхи, Л., 1970; Smith G. M., Cryptogamic botany, 2ed., v. 2, N. Y., 1955; S с h u s t e r R. M., The Hepaticae and Anthocerotae of North America, v. 1, N. Y.- L., 1966; P a r i-h a r N. S., An introduction to embryophyta, 5 ed., v. 1, Allahabad, 1969. И. И. Абрамов.
МХИТАР ГОШ [1120(?), Гандзак,- 1213, с. Нор-Гетик, ныне Гош], армянский мыслитель, литературный и общественный деятель. Автор более десяти трудов, среди которых большую ценность представляют "Басни" (изд. 1790) и первый арм. судебник (см. Мхитара Гоша судебник). M. Г.- один из ранних провозвестников арм. Возрождения. Темы его басен социальны, что было новшеством для того времени; автор прославляет добро, труд, разоблачает пороки. Басни средневековой Армении, пер. и предисл. И. А. Орбели, М.- Л., 1956. Лит.: M а р р H. Я., Сборники притч Вардана, ч. 1, СПБ, 1899. МХИТАРА ГОША СУДЕБНИК, юридич. сборник, составленный в 1184 Мхитаром Гошем. Одним из гл. источников M. Г. с. было арм. обычное право; кроме того, в него был включён ряд положений, заимствованных из визант. права, церк. постановлений и др. M. Г. с. состоит из введения и 2 частей: церк. каноны и светские законы. К.-л. строгой системы в судебнике нет, теоретич. рассуждения сочетаются с предложениями составителя, нормами права и их толкованием. M. Г. с. был направлен на защиту феод, отношений, однако в нём имеется ряд наставлений морального характера об ограничении произвола феодалов. Хотя M. Г. с. не носил офиц. характера, он применялся как в самой Армении, так и за её пределами, где имелось арм. население. Лит.: Армянский судебник Мхитара
Гоша, пер. с древнеармянского, [Ep., 1954].
МХИТАPИCTЫ, конгрегация армянских
церковников, осн. в 1701 монахом Мхи-таром Себастаци, с 1717 - ок. Венеции
на о. Сан-Ладзаро, где существует и поныне. Позже часть организации обосновалась
в Вене, где также продолжает существовать. M. сыграли известную роль в
развитии арм. филологии и церковно-феод. лит-ры. Они издавали труды др.-арм.
писателей, исследования по армянове-дению, словари, переводили лит. памятники
др.-греч. и рим. лит-ры на др.-арм. яз. Деятельность M. в основном служила
укреплению влияния Ватикана на Бл. Востоке; наносит большой вред социальной
и нац.-освободит, борьбе зарубежных армян.
МЦЕНСК, город областного подчинения, центр Мценского р-на Орловской обл. РСФСР. Расположен на р. Зуша (приток Оки). Ж.-д. станция на линии Орёл - Тула, в 49 км к С.-В. от Орла. Через город проходит автомагистраль Москва - Симферополь. 28 тыс. жит. (1970). Впервые упоминается в 1147 под назв. Мьченск в составе Черниговского княжества. В 1320 присоединён к Литве; с нач. 16 в.- в составе Рус. гос-ва. С 1778- уездный город. 3-ды: вторичных цветных металлов, алюминиевого литья (филиал Моск. автозавода им. Лихачёва), коммунального машиностроения, кормовых антибиотиков, кирпичный; овощеконсервный, мясной комбинаты; швейная, мебельная ф-ки. Крупный цветоводческий совхоз. Вечерний металлургич. техникум. В 10 км к С. от M.- заповедник-усадьба И. С. Тургенева Спасское-Лутовиново. Лит.: Макашов А., Ровесник Москвы,
Орел, 1972.
МЦХЕТА, город, центр Мцхетского р-на Груз. CCP. Расположен при впадении р. Арагви в р. Кура, на Воен.-Грузинской дороге. Ж.-д. станция на линии Тбилиси - Самтредиа, в 21 км к С. от Тбилиси. 7,1 тыс. жит. (1970). M.- древний город Грузии. Осн. во 2-й пол. 1-го тыс. до н. э. До кон. 5 в. н. э. M.- столица вост.-груз, гос-ва Картли (Иберии). В период средневековья M. оставалась значит, городским, торгово-ремесл. и особенно религ. центром (резиденция главы груз, церкви - католикоса). Раскопками в 1870-х гг. и особенно с 1937 в M. вскрыты остатки укреплённых резиденций, гор. кварталов, а также могильники, датируемые от энеолита до позднего средневековья. К эпохам бронзы и раннего железа (2-1-е тыс. до н. э.) относятся могильники (см., напр., Мцхета. Остатки дворцового сооружения с колоннами в Армазцихе (Багинети). Самтаврский могильник), связанные с поселениями оседлых земледельцев, занимавшихся и скотоводством. На базе этих поселений возник крупный благоустроенный антич. город. На прав, берегу Куры открыты раскопками развалины акрополя Армазцихе (2-я пол. 1-го тыс. до н. э.- нач. 1-го тыс. н. э.). К позднеантич. времени относится исследованный в 1951 склеп кон. 1- нач. 2 вв. н. э.- прямоугольное сооружение (2,3 X X 1,8 X 1,9 м) из рваного камня с полуциркульным сводом из тёсаных кам. блоков и двускатным перекрытием из укреплённой известковым раствором крупной черепицы. Внутри - стены, свод и пол, снаружи - вост. фасад облицованы хорошо отёсанными и пригнанными квад-рами песчаника. Дверь была заложена массивной кам. плитой. В склепе обнаружены кости женщины и ребёнка, доски и металлич. детали от саркофага и остатки богатого инвентаря: золотая пряжка с камнями, десятки золотых бус, подвесок и пуговиц; кам. и пастовые бусы и вставки; бронзовая литая фигурка поющего юноши; разноцветные стеклянные тарелочки и бальзамарии; керамика; 69 серебряных монет (1 в. до н. э. и 1 в. н. э.) и 10 золотых (4 рим. ауреуса, 5 груз, подражаний статерам и одна заготовка для такового). Мцхета: 1 - остатки грунтового погребения знатного воина из Самтаврского могильника. Нач. 1-го тыс. до н. э.; 2 - гробница питиахша Ас-пурга (2 в. н. э.) и женщины (3 в. н. э.) В фамильной усыпальнице питиахшей. Склеп находится в окружении небогатых черепичных гробниц 1-2 вв. н. э. Рядом с ним обнаружен скелет лошади. Выше по течению Куры в Армазисхеви обнаружены развалины дворцового комплекса, баня (2-3 вв.) и некрополь (первые века н. э.). Среди ср.-век. памятников M.- комплекс монастыря Самтавро (главный храм к-рого - крестово-купольное здание 11 в. с богатым скульпт. декором на фасадах), кафедральный собор Светицховели. Близ M.- храм Джвари, Земо-Авчальская ГЭС им. В. И. Ленина (ЗАГЭС; 1927, арх. A. H. Кальгин, К. А. Леонтьев, M. С. Мачавариани) с пам. В. И. Ленину (бронза, камень, открыт в 1927, скульптор И. Д. Шадр). Илл. см. т. 2, стр. 298; т. 7, табл. XXXIV, XXXVI (стр. 384-385) и стр. 385. Лит.: Северов H. П., Ч у б и-нашвили
Г. H., Мцхета, M., 1946; Ц и ц и ш в и л и И. H., Гробница у станции Мцхета,
"Краткие сообщения о докладах и полевых исследованиях Института истории
материальной культуры АН СССР, 1954, в. 54; Ломтатпдзе Г. А., Археологические
раскопки в древнегрузинской столице Мцхета, Тб., 1955; Мцхета. Итоги археологических
исследований, т. 1 - Археологические памятники Армазис-Хеви по раскопкам
1937-1946 гг., Тб., 1958; Г в е р дц и т е л н Р., Мцхета, Тб., 1962; [Ч
ан и ш в и л и Г. К.], Мцхета, Тб., 1965.
M-ЧИСЛО, отношение скорости "МШАК" ("Труженик"), армянская литературно-политическая
газета. Издавалась в 1872-1920 в Тбилиси. Была основана либеральным публицистом
и критиком Г. Арцруни. После его смерти в 1892 "M." редактировали
А. Калантар, А. Аракелян и др. Газета выступала с ли-берально-бурж. позиций
против реакц. духовенства, косности консерваторов, писала о тяжёлом существовании
армян в Турции, ставила вопросы просвещения народа, эмансипации женщин,
развития арм. лит-ры и иск-ва. Угнетённое положение крестьян и рабочих
также находило отражение на страницах "М.". В газете печатались произв.
Раффи, P. Патканяна, А. Ширванзаде и др. В 1917-20 "М." стала рупором бурж.
национализма.
MШAHKA (Sagina), род растений сем.
гвоздичных. Многолетние или однолетние травы, часто образующие дерновинки.
Стебли тонкие, лежачие или приподнимающиеся, с линейно-ланцетными или шиловидными
листьями. Цветки б. ч. мелкие, одиночные пазушные или в соцветиях. Околоцветник
5-членный; лепестки обычно белые, иногда их нет. Плод - коробочка. Ок.
30 видов, гл. обр. в умеренном поясе Сев. полушария, а также в горах Юж.
и Вост. Африки, Юж. Америки и о. H. Гвинея. В СССР 8 видов. Широко распространены
M. лежачая (S. procumbens) и M. узловатая (S. nodosa) - обычные растения
на сырых лугах, берегах водоёмов, выгонах, у дорог. M. моховидная (S. saginoi-des)
растет в арктич. зоне и альп. поясе гор по каменистым склонам, скалам,
галечникам; служит кормом для овец.
МШАНКИ (Bryozoa, или Polyzoa), класс беспозвоночных животных типа щупальцевых (Tentaculata). Водные, преим. морские, сидячие, колониальные животные. Одни колонии M. обрастают субстрат (камни, водоросли и пр.) в виде корочек и комков (рис. 1,1), другие - древовидные, листовидные, кустикообразные (рис. 1, 2), третьи - имеют вид студенистых тяжей. Колонии M. состоят из микроскопич. особей - зооидов, заключённых в известковую, хитиноидную или студенистую ячейку (цистид) (рис. 2). Через отверстие цистида может выдвигаться передняя часть тела (полипид), несущая ротовое отверстие с венчиком щупалец. Движение ресничек на щупальцах создает ток воды, к-рым ко рту приносятся мелкий планктон и детрит - осн. пища M. Рис. 1. Мшанки 1 - Flustrella hispida;2 - Dendrobeania flustroides; 3 - Bugula tricuspis. В связи с сидячим образом жизни все системы органов упрощены. Рот ведет в глотку, далее следует пищевод, желудок, петлеобразно изогнутая кишка. Анальное отверстие лежит на спинной стороне тела близ рта, но вне венчика щупалец. Дыхательная, кровеносная и выделительная системы у большинства M. отсутствуют. Дыхание осуществляется через щупальца и поверхность тела. Имеется вторичная полость, жидкость к-рой выполняет функции крови. Выделяемые вещества собираются в фагоцитах и выводятся через кишечник. Нервная система состоит из ганглия и отходящих от него нервов. Мышцы продольные, втягивающие полипид, и париетальные, выпячивающие его. Размножение бесполое и половое. Большинство M. двуполы. Яйца развиваются в воде, в полости тела или в выводковых камерах, овицел-лах, гонозооидах и др Личинки типа трохофоры различной формы, снабжены венчиком ресничек, а нек-рые (цифонаутес) - и двустворчатой раковиной. Рис 2. Схема двух зооидов а - со втянутым полипидом; 6 - с высунувшимся полипидом. Осев на дно, личинка прикрепляется и, претерпев ряд упрощений, образует первую особь - анцеструлу, к-рая почкованием дает начало новой колонии. У MH. пресноводных M. развиваются внутр. почки, т. н. статобласты, к-рые перезимовы-вают и весной образуют новые колонии. В колониях многих мор. M. набчю-дается полиморфизм, т. е. наличие особей, отличных по строению и функции от нормальных аутозооидов. Среди них различают гонозооиды, авикулярии, вибра-кулярии (служащие для защиты и очистки колонии), кенозооиды, укрепляющие колонию. Для M. характерны недолговечность отд. особей, быстрое отмирание и дегенерация всего зооида или его части с последующим восстановлением. M. широко распространены в пресных водах и морях, где обитают от полосы прилива до глуб. 200-300 м (реже до 6000м). Ок. 4500 видов; в морях СССР - ок. 450 видов, в пресных водах - ок. 30 видов. Наиболее древние остатки M. известны из нижнего ордовика, но предполагают, что они существовали уже в кембрии. Палеонтологи выделяют M. в особый тип Bryozoa с 2 классами· Gym-nolaemata (голоротых) и Phylactolaemata (покрыторотых); последние в ископаемом состоянии почти неизвестны. Всего ок. 15 000 вымерших видов. Наибольшего разнообразия достигали в палеозое. В ряде случаев M. принимали участие в образовании рифов. Имеют значение для решения вопросов стратиграфии, палеоэкологии и палеогеографии. Лит . К л ю г е Г. А., Мшанки (Bryozoa),
в кн.: Жизнь пресных вод СССР, под ред. В И Жадина, т 2, M - Л., 1949;
его же, Мшанки северных морей СССР, M - Л , 1962 (Определители по фауне
СССР, т. 76); Основы палеонтологии, под ред T. Г. Сарычевой, т 7, M , 1960;
The Lophophorate Coelomates-Pbylum Ectoprocta, в KH Hyman L H, The Invertebrates, E И. Андросова.
MШАТТА, замок династии Омейядов в Иордании (8 в.; по мнению некоторых исследователей, 3-4 вв.), в 25 км к Ю.-В. от Аммана; не был окончен, ныне - руины. Представлял собой квадратное в плане сооружение (с регулярной застройкой по сторонам внутреннего двора), окруженное стенами с башнями. Прославился орнаментальным фризом, вырезанным в камне, в нижней части юж. оборонительной стены. Фриз состоит из крупных треугольников, заполненных реалистически трактованными ювелирно-тонкими узорами из переплетенных виноградных лоз, пальметт, розеток, листьев аканфа, цветов и животных. Лит : Trumpelmann L., Mschatta,
Tubingen, 1962.
МШВЕЛИДЗЕ Шалва Михайлович [р. 15(28)5.1904, Тбилиси], советский композитор, нар. арт. Груз. CCP (1958). Чл. КПСС с 1947. Окончил в 1930 Тбилисскую консерваторию по классу композиции у M. M. Багриновского, учился в аспирантуре Ленингр. и Тбилисской консерваторий у В. В. Щербачева. Автор опер "Сказание о Тариэле" (1946) и "Десница великого мастера" (1961)- обе поставлены в Тбилисском театре оперы и балета им. 3. Палиашвили; ораторий "Кавка-сиони" (1949) и "Векам в предание" (1970, поев. В. И. Ленину), 4 симфоний (1943, 1944, 1952, 1968), симфонич. поэм "Звиа-даури" (1940), "Миндия" (1950), "Юноша и барс" (1962), симфонич. сюит, увертюр, сб. песен, музыки к драматич. спектаклям и фильмам. Преподаёт в Тбилисской консерватории (с 1942 проф.). В 1941-52 пред. Союза композиторов Грузии. Деп. Верх. Совета Груз. CCP 3-го созыва. Гос. пр. СССР (1942, 1947). Гос. пр. Груз. CCP им. 3. Палиашвили (1971). Инд. пр. им. Джавахарлала Неру за муз. произв. на инд. темы и серию очерков о муз. фочьклоре Индии (1973). Награжден орденом Ленина, 2 др. орденами, а также медалями. MЫЙЗАКЮЛА, город в Вильяндиском
р-не Эст. CCP, у границы с Латв. CCP. Ж.-д. линии на Пярну, Тюри, Руиену.
Цех Талчинского экскаваторного з-да, льнопрядильная ф-ка, ремонт ж.-д.
контейнеров.
MЫЛA, река в Коми АССР, прав, приток р. Цильма (басе. Печоры). Дч. 186 км, пл. басе. 2700 км2. Берет начало с Тиман-ского кряжа. Питание преим. снеговое. Замерзает в ноябре, вскрывается в начале мая. Сплавная. Мшатга. Деталь рельефного фриза.
МЫЛА, соли высших жирных кислот (см. Карбоновые кислоты). В произ-ве и быту M. (или т оварными M.) наз. технич. смеси водорастворимых солей этих K-T, часто с добавками нек-рых др. веществ, обладающие моющим действием. Основу смесей обычно составляют натриевые (реже калиевые и аммониевые) соли насыщенных и ненасыщенных жирных K-T с числом атомов углерода в молекуле от 12 до 18 (стеариновой, пальмитиновой, миристиновой, лауриновой и олеиновой). К M. часто относят также соли нафтеновых и смоляных K-T, а иногда и др. соединения, обладающие в растворах моющей способностью. Не растворяющиеся в воде соли жирных K-T и щёлочноземельных, а также поливалентных металлов наз. "металлическими" M. Водорастворимые M.- типичные мицел-лообразующие поверхностно-активные вещества. При концентрации выше определённого критич. значения в мыльном растворе наряду с отдельными молекулами (ионами) растворённого вещества находятся мицеллы - коллоидные частицы, образованные скоплением молекул в крупные ассоциаты (см. также Полуколлоидные системы). Наличие мицелл и высокая поверхностная (адсорбционная) активность M. обусловливают характерные свойства мыльных растворов: способность отмывать загрязнения, пениться, смачивать гидрофобные поверхности, эмульгировать масла и др. Приготовление M. обработкой жиров растит,
золой, известью и естеств. щелочами, по свидетельству Плиния Старшего,
было известно ещё древним галлам и германцам. Упоминание о M. встречается
у римского врача Галена (2 в. н. э.). Однако как моющее средство
M.
стали использовать значительно позже; к 17 в. оно, по-видимому, было уже
достаточно распространено в Европе. Мыловаренная пром-сть возникла в 19
в., чему способствовали развитие химии жиров (работы франц. химика M. Э.
Шеврёля, 1813- 1823) и создание достаточно широкого произ-ва соды по способу
франц. химика H. Леблана (1820). Совр. мыловаренная пром-сть выпускает
M. различных типов и сортов. По назначению различают Технологич. процесс получения M. складывается из 2 этапов: варки M. и переработки сваренного M. в товарный продукт. Варку M. проводят в спец. аппаратах - варочных котлах. Жировое сырьё при нагревании подвергают омылению едкой щёлочью, обычно каустич. содой (гидроокисью натрия); при этом жиры превращаются в смесь солей жирных K-T и глицерин. Иногда используют жиры, предварительно подвергнутые гидролизу (расщеплению) с образованием свободных жирных K-T. Расщеплённые жиры в варочном котле нейтрализуют кальцинированной содой (карбонатом натрия), а затем доомыляют едкой щёлочью. В обоих случаях в результате варки образуется мыльный клей - однородная вязкая жидкость, густеющая при охлаждении. Товарное M., полученное непосредственно из мыльного клея, наз. клеевым; содержание жирных K-T в нём обычно находится в пределах от 40 до 60% . Обработка мыльного клея электролитами (отсолка) вызывает его расслоение. При полной отсолке растворами едкой щёлочи или хлористого натрия в варочном котле возникают два слоя. Верх, слой - концентрированный раствор M., содержащий не менее 60% жирных кислот, наз. мыльным я д-р о м. Из него получают товарное M. высших сортов (ядровое M.). Нижний слой - раствор электролита с малым содержанием M.- подмыльный щёлок; в него переходит большая часть глицерина (к-рый извлекают как ценный побочный продукт произ-ва) и загрязнений, внесённых в мыльный клей с исходными продуктами. Метод получения клеевых M. принято наз. прямым, ядровых - косвенным. В произ-ве хозяйственных M. используют оба эти метода. Туалетные M., как правило, готовят косвенным методом, причём мыльное ядро получают из лучшего жирового сырья и подвергают дополнит, очистке. На втором этапе при получении твёрдых M. мыльную массу - продукт варки - охлаждают, подсушивают, а затем механич. обработкой с помощью спец. аппаратуры придают ей пластичность и однородность, формуют и разрезают на куски стандартной массы. В туалетные M. вводят отдушки, красители, антиокси-данты, а в нек-рых случаях - дезинфицирующие, лечебно-профилактические, пенообразующие и др. специфичные добавки. В дешёвые сорта M. иногда добавляют минеральные наполнители - бентонитовые глины, очищенный каолин. Особую группу составляют пережиренные туалетные мыла; в них отсутствует свободная щёлочь и обычно содержатся косметич. добавки (высшие жирные спирты, питат. вещества и др.). См. также Косметика. Порошкообразные M. получают распылительной сушкой мыльных растворов. В продажу они поступают без добавок (мыльные порошки) или в смеси со значит, количеством щелочных электролитов (содой, фосфатами и др.), к-рые улучшают моющую способность M. (стиральные порошки). При производстве M. применяется автоматизированная технологич. аппаратура непрерывного действия. Мировое произ-во хозяйственных M. постепенно сокращается в связи с увеличением выпуска синтетич. моющих средств и растущим дефицитом жирового сырья. Однако с распространением разнообразных синтетич. мылоподобных веществ M. не потеряли своего значения важнейшего средства личной гигиены. Они по-прежнему широко применяются в быту и во многих отраслях пром-сти (особенно в текстильной). M. наряду с др. типами поверхностно-активных веществ используются как смачиватели, эмульгаторы, стабилизаторы коллоидно-дисперсных систем. M. применяют в составе смазочно-охлаждающих жидкостей для металло-обр. станков; при обогащении полезных ископаемых флотацией. Их используют в хим. технологии: при синтезе полимеров эмульсионным способом, в произ-ве лакокрасочной продукции и др. "Металлические" M. как загустители входят в состав пластичных смазок, как сиккативы (ускорители чвысыхания") - в состав масляных лаков, олиф и др. Лит.: Технология переработки жиров, 3 изд., M., 1963; Справочник по мыловаренному производству, под ред. И. M. Товбина, M., 1974. Л. А. Шиц. МЫЛО ЖИДКОЕ, 1) туалетное M. ж.- прозрачный водно-спиртовой раствор калийного жирового мыла; содержит до 20% жирных K-T, 15-20% этилового спирта и до 1% отдушки. Применяется гл. обр. для мытья головы и рук. 2) Хозяйственное M. ж.- жидкий или мазеобразный продукт омыления расщеплённых жирных масел растительных; содержит 40 - 60% жирных K-T; сырьём для получения хозяйственного M. ж. служат также отходы жироперера-батывающих произ-в, нафтеновые к-ты, канифоль и талловое масло. Предназначено для стирки технич. тканей и текст, изделий, мойки шерсти и т. п. MЫЛО ЗЕЛЁНОЕ, дезинфицирующее средство. Получают M. з. омылением растит, масел раствором едкого кали. Применяют для очищения кожи и приготовления мыльного спирта и мыльно-карболового раствора, употребляемых в качестве дезинфекционных средств. Входит в состав Вилъкинсона мази. МЫЛО СУЛЬФАТНОЕ, побочный продукт произ-ва целлюлозы сульфатным способом. Образуется при варке смолистой (кедровой, сосновой) древесины в щелочном растворе. Содержит до 55% жирных и смоляных K-T. M. с. перерабатывают на талловое масло, применяемое в лакокрасочной пром-сти и произ-ве технич. поверхностно-активных веществ. МЫЛОНАФТ, технич. смесь натриевых солей нафтеновых K-T, получаемая как отход при щелочной очистке керосиновых, газойлевых и соляровых дистиллятов нефти. M. - мазеобразный продукт от соломенно-жёлтого до тёмно-коричневого цвета с неприятным запахом; содержит не менее 43% нафтенатов (в пересчёте на кислоты), 9-15% неомыляемых веществ, 4-6% минеральных солей, а также воду. Асидол-M, содержит также свободные нафтеновые к-ты; его получают обработкой M. серной к-той. M. и асидол-М. применяют в качестве эмульгаторов, заменителей жиров в мыловаренном произ-ве. M. иногда используют как моющее и дезинфицирующее средство. Лит.: Химические товары, 3 изд.. т. 2, M., 1969, с. 425, 463. МЫЛЬНАЯ КОРА, панамская кора, кора дерева Quilliaja saponaria (сем. розовых) родом из Юж. Америки. Содержит до 9% сапонинов; используется как вспенивающее средство при изготовлении эмульсий, а также вместо мыла при стирке шёлковых и шерстяных тканей. МЫЛЬНИКОВ Андрей Андреевич (р. 22.2.1919, Покровск, ныне Энгельс Саратовской обл.), советский живописец, нар. худ. РСФСР (1968), действит. чл. AX СССР (1966). Учился в Ленинграде в Институте живописи, скульптуры и архитектуры (1938-46) у И. Э. Грабаря и В. M. Орешникова. С 1947 преподаёт там же (с 1967 проф.). Работы M. (жанровые картины, пейзажи, портреты, мозаичные панно, монументально-декоративные росписи) отличаются эмоциональной приподнятостью, ясностью композиции, праздничной яркостью цвета. Произв.: "На мирных полях" (1949; Гос. пр. СССР, 1951) и "Пробуждение" (1957) - оба в Русском музее, Ленинград; "Утро" (1972, Дирекция художественных фондов и проектирования памятников Мин-ва культуры РСФСР); "Ленин" (1961) - жёсткий занавес Кремлёвского Дворца съездов; мозаики в Ленинградском ТЮЗе (1962). Награждён орденом "Знак Почёта" и медалями. Лит.: [Б л э к В. Б.], А. А. Мыльников. Альбом, Л., 1960. МЫЛЬНИКОВ Григорий Михайлович (р.
25.5.1919, с. Егорьевка, ныне Ka-сторненского р-на Курской обл.), майор
(1944), дважды Герой Сов. Союза (23. 2.1945 и 19.4.1945). Чл. КПСС с 1942.
В Сов. Армии с 1939. Окончил Борисоглебскую военную школу лётчиков (1940).
В Великую Отечеств, войну 1941-45-на Западном, Ленинградском и 3-м Белорусском
фронтах. Был пилотом, командиром звена, зам. командира эскадрильи, командиром
эскадрильи 15-го гвардейского штурмового авиаполка и зам. командира полка.
Произвёл 223 боевых вылета. С ноября 1945 по состоянию здоровья в запасе.
Награждён орденом Ленина, 3 орденами Красного Знамени, орденами Отечественной
войны 1-й степени, Александра Невского, Красной Звезды и медалями. Портрет
стр. 154.
МЫЛЬНОЕ ДЕРЕВО (Sapindus), род растений сем. сапиндовых. Вечнозелёные или листопадные деревья с очередными перистыми листьями. Цветки в конечных или пазушных метёлках. Ок. 15 видов в тропиках Азии и Америки. Плоды M. д. содержат до 38% сапонинов и используются вместо мыла. Культивируется в тропич. и тёплых странах как декоративное и для получения сапонинов. В СССР на Черноморском побережье Кавказа и изредка в Закавказье и Cp. Азии выращивают 3 вида: M. д. настоящее (S. sa-ponaria), M. д. Мукоросси, или китайское (S. mucorossii), и зимостойкий северо-амер. вид - M. д. Друммонда (S. drum-mondii). Иногда M. д. наз. к е л ь р е ут е р и ю (Koelreuteria paniculata) - дерево того же семейства, растущее в Восточной Азии и содержащее сапонины в коре. Лит.: Деревья и кустарники СССР,
т. 4, М.- Л., 1958.
МЫЛЬНЫЙ КОРЕНЬ, корневища и корни растений, содержащие в значит, количестве сапонины - органич. вещества, придающие растворам способность пениться. Из M. к. получают мед, и тех-нич. сапонины; отвары M. к. используют в произ-ве жидких инсектицидов, применяют как отхаркивающее средство. Раньше M. к. использовали для стирки тканей вместо мыла (отсюда назв.). M. к. получают от растений разных семейств, в СССР - преимущественно семейство гвоздичных, например мыльнянка лекарственная даёт "красный M. к." (содержащий 13-15% сапонинов; по др. данным, до 35% ), качим метельчатый -"белый M. к." (6,5-20% сапонинов), колюче-листник железистый и др. его виды - "туркестанский M. к.". МЫЛЬНЯНКА, сапонария (Saponaria), род травянистых растений сем. гвоздичных. Цветки в метельчатых или головчатых соцветиях; чашечка спайно-листная; венчик розовый, белый, иногда жёлтый, из 5 лепестков. Плод - одно-гнёздная коробочка. Ок. 30 видов в Евразии, гл. обр. в Средиземноморье, на В.- до Зап. Сибири и Cp. Азии. В СССР 10 видов, в т. ч. M. лекарственная (S. officinalis), растущая по опушкам лесов,в долинах рек; богатые сапонинами корневища и корни её используют в пром-сти под названием мыльного корня красного, а также в медицине (преимущественно как отхаркивающее); её махровые формы применяют в цветоводстве. Мыльнянка лекарственная.
МЫНАРАЛ, посёлок гор. типа в Мойын-кумском
р-не Джамбулской обл. Казах. CCP. Расположен на юго-зап. берегу оз. Балхаш,
в 4 км от ж.-д. станции Мынарал (на линии Моинты - Чу). Рыбозавод.
МЫРЗАБАЕВ Артык (р. 11.5.1930, Кызыласкерский
район, ныне в черте города Фрунзе), киргизский советский певец (баритон),
нар. арт. СССР (1967). В 1954 окончил Муз. уч-ще им. Куренке-ева в г. Фрунзе.
С того же года солист Кирг. театра оперы и балета. Обладает голосом красивого
тембра, развитой вокальной техникой, сценич. дарованием. Лучшие партии:
Искандер, Токтогул ("Аджал орду на" и "Токтогул" Власова, Малдыбаева и
Фере), Артём ("Ак шум-кар" Ряузова), Фигаро ("Севильский цирюльник" Россини),
Яго ("Отелло" Верди), Онегин ("Евгений Онегин" Чайковского) и др. Выступал
на Всемирной выставке в Монреале (1967). Гастролировал в Китае, Монголии,
Румынии и др. странах.
МЫРКВИЧКА (Мърквичка) Иван (23.4.1856, Видим, Чехия,-16.5.1938, Прага), болгарский живописец и график. По национальности чех. Учился в пражской (1873-76) и мюнхенской (1876- 1877) AX. В 1881-1921 жил в Болгарии. Один из создателей Гос. рисовального училища в Софии (преподаватель и директор в 1896-1921). Чл. Болг. АН (1918). Представитель демократич. реализма, M. изображал гл. обр. нар. празднества, сцены крест, труда, передавал своеобразие болг. нац. обычаев и костюмов ("Базар в Пловдиве", 1888; "Рученица", илл. см. т. 3, стр. 497; "Беженцы из Фракии", 1897; все - в Нац. художеств, гал., София). Известен также как портретист и иллюстратор. Лит: Львова Е. Иван Мырквичка, M.,
1955.
MЫC, участок побережья, вдающийся в море, озеро или реку. Может быть сложен как коренными породами, так и наносами. В последнем случае M. обычно низменны и представляют собой оконечности береговых аккумулятивных форм. МЫС ШМИДТА, посёлок гор. типа, центр
Шмидтовского р-на Чукотского нац. округа Магаданской обл. РСФСР. Расположен
на берегу Чукотского моря. Ионосферная станция. Радиометеорологич. центр.
Назван в честь О. Ю. Шмидта.
MЫСИК, Мысык Василий Александрович [р. 11(24).7.1907, с. Новопавловка, ныне Межевского р-на Днепропетровской обл.], украинский советский поэт. Род. в крест, семье. Учился на физико-мате-матич. ф-те Харьковского ун-та (1956- 1957). Участник Великой Отечеств, войны 1941-45. Первые стихи опубл. в 1923; в 1927 вышел сб. "Травы", отмеченный яркой образной манерой, тяготеющей к, классичности форм и филос. осмыслению действительности. Поездка в республики Cp. Азии обогатила поэзию M. новыми ритмами и настроениями (сб-ки "Четыре ветра", 1931; "Строители", 1933; "Турк-сио", 1933). После большого перерыва в лит. деятельности опубл. сб-ки "Борозды" (1962), "Вершины" (1963), "Чернотропз" (1966, рус. пер. 1967), "Поле" (1970), в к-рых он настойчиво возвращается к разговору о личной ответственности человека за мир на земле, о необходимости помнить уроки истории и бережно хранить традиции своего народа. Перевёл на укр. яз. стихи P. Бёрнса, Дж. Кит-са, P. Тагора, Рудаки, Омара Хайяма, Сзади. Соч.: Вибране, К., 1958; Берег, К., 1972; в рус. пер.- Избранное. [Предисл. T. Гнедич], M., 1973. С. П. Князева.
МЫСКИ, город (до 1956-посёлок) областного
подчинения в Кемеровской обл. РСФСР. Расположен в Горной HIo-рии, при впадении
р. Мрас-Су в Томь. Ж.-д. станция в 60 км к В. от Новокузнецка. 36,2
тыс. жит. (1970). Предприятия деревообрабатывающей промышленности. Близ
M.- Томь-Усинская ГРЭС. Добыча угля.
МЫСЛИВЕЧЕК (Myslivecek) Йосеф (9.3.1737, Прага,-4.2.1781, Рим), чешский композитор и дирижёр. Чл. Болон-ской филармонич. академии (1771). Учился в Праге, где создал первые симфонии (1762). С 1763 жил в Италии. Первая опера M.- "Медея" (Парма, 1764); получил известность после пост. "Бел-лерофонта" (Неаполь, 1767). В 1770 ? Болонье M. встретился с юным В. А. Моцартом, сразу же высоко оценившим его музыку; M. был дружен с ним много лет. M.- автор ок. 30 опер (шли с огромным успехом в 1767-80 в различных городах Италии, а также в Праге и Вене), в к-рых развивал традиции итал. оперы-сериа (см. Опера). Среди лучших- "Гипермнестра", "Тамерлан Великий", "Монтесума", "Эцио", "Олимпиада", "Армида", "Медонт, царь Эпир-ский". Автор ораторий, в т. ч. "Авраам и Исаак"; симфоний, увертюр, нескольких концертов для струнных инструментов с оркестром, струнных ансамблей, трио-сонат, соч. для чембало и др. Музыка M., испытавшего итальянские влияния, содержит элементы нар. чеш. песенно-сти. Как дирижёр пропагандировал оперное творчество К. В. Глюка. Лит.: Б э л з а И., Очерки развития чешской музыкальной классики, M.- Л., 1951, с. 82-89; его же. История чешской музыкальной культуры, т. M., 1959, с. 285- 306; Шагинян M., Йозеф Мысливечек, Г. М. Мыльников. В. И. Мыхлик. 2 изд., M., 1968; CeI eda J., Josef My-slivecek..., Praha, 1946 (с библиографич. указателем). И. Ф. Бэлза. МЫСЛОВИЦЕ (Mystowice), город в Польше,
в Катовицком воеводстве, в пределах Верхнесилезской агломерации. 46 тыс.
жит. (1972). Ж.-д. узел. Добыча кам. угля; произ-во шахтного оборудования,
огнеупоров, электротехнич. фарфора.
"МЫСЛЬ", большевистский легальный ежемесячный филос. и общественно-экономич. журнал. Издавался в Москве с дек. 1910 по апр. 1911; вышло 5 номеров. Офиц. редактором-издателем "М." был П. К. Пирожков, фактич. редактором - В. И. Ленин, руководивший журналом из-за границы. Активно участвовали В. В. Боровский, И. И. Скворцов-Степанов, M. С. Ольминский и др. В "М." сотрудничали Г. В. Плеханов, меньшевики-партийцы. В журнале было опубликовано 6 статей Ленина. Последний, 5-й номер был конфискован, и "М." закрыта. Продолжением её явился журн. "Просвещение", выходивший в Петербурге. Лит : Большевистская периодическая
печать (декабрь 1900 - октябрь 1917). Библиографический указатель, M.,
1964, с. 105.
"МЫСЛЬ", издательство социально-экономич. литературы в системе Гос. к-та Сов. Мин. СССР по делам издательств, полиграфии и книжной торговли. Находится в Москве. Образовано в 1930 как Изд-во социально-экономич. лит-ры - Соцэкгиз (в результате реорганизации социально-экономич. отдела Госиздата, основанного в 1919); в 1941 Соцэкгиз был слит с Госполитиздатом, в 1957 вновь выделен в самостоят, изд-во, в 1963 к Соц-экгизу присоединены Издательство Высшей парт, школы и Академии обществ, наук при ЦК КПСС и Географгиз; образованное изд-во получило совр. название. Выпускает исследования, монографии, на уч.-популярные книги по марксистско-ленинской философии, истории философии, науч. атеизму, экономике социализма и коммунизма, мировой экономике, истории СССР и всеобщей истории, учебные и методич. пособия по обществ, наукам для высших партийных уч. заведений и системы политич. просвещения, по экономич. и физич. географии СССР и зарубежных стран. В 1973 в составе изд-ва имелись 3 специализированные главные редакции: социально-экономич. лит-ры; учебно-методич. лит-ры для парт. уч. заведений; географич. лит-ры. В 1972 объём издательской продукции составил 292 названия (175,8 млн. печатных листов-оттисков) тиражом св. 14 млн. экз. А. П. Порываев. МЫСЛЬБЕК (Myslbek) Йосеф Вацлав (20.6.1848, Прага,-2.6.1922, там же), чешский скульптор. Учился в мастерской В. Левы и в AX в Праге. Проф. Уч-ща декоративных иск-в (1885-96) и AX (1896-1919) в Праге. Создавал возвышенные, монументальные образы ист. деятелей Чехии и героев народного эпоса, сочетающие символич. обобщённость с жизненной конкретностью (проект пам. Я. Жижке для г. Табор, бронза, 1870-е гг.; скульпт. группы на мосту Па-лацкого в Праге, бронза, 1881-97). Наиболее значит, произв. M.- пам. св. Вацлаву в Праге (бронза; открыт в 1913). Автор многочисл. портретов Лит.: S t е с h V. V-, J. V. Myslbek,
Praha, 1952.
МЫСХАКО, мыс южнее Новороссийска, на к-ром во время Великой Отечеств, войны 1941-45 в ночь на 4 февр. 1943 в р-не Станички корабли Черноморского флота высадили отряд морской пехоты числ. 800 чел. под командованием майора Ц. Л. Куникова. Отряд захватил плацдарм, названный в дальнейшем "Малой землёй", к-рый, несмотря на яростные атаки нем.-фаш. войск с суши и с воздуха, остался в руках сов. войск. 6 февр. отряд получил подкрепление и ему удалось не только удержать плацдарм, но и расширить его. Плацдарм у M. сыграл важную роль в Новороссии-ско-Таманской операции 1943. МЫТ, одна из древнейших рус. пошлин с товаров и лиц, а также место её сбора. Упоминается в документах с 12 в. С сер. 13 в. M. взимался при провозе товаров через заставы у городов и крупных селений. Различался "сухой" и "водяной" M. Право обложения M. принадлежало гос-ву, но его сбор нередко передавался феодалам. M. дал начало появлению различных проезжих торг, пошлин, большинство к-рых было упразднено Торговым уставом 1653. В 1653-1753 под M. понималась 5%-ная пошлина, взимавшаяся в Москве и Ниж. Новгороде с леса, сена, рогатого скота, съестных припасов (кроме хлеба) и др. на т. н. Мытных дворах. МЫТ, острая инфекционная болезнь лошадей, проявляющаяся гнойно-катаральным воспалением слизистой оболочки носоглотки и подчелюстных лимфатич. узлов. Распространён повсеместно, чаще - в странах с умеренным и холодным климатом; в СССР регистрируется спорадически. Возбудитель M.- мытный стрептококк (Streptococcus equi), обладает значит, устойчивостью во внеш. среде. Источник возбудителя инфекции - больные и переболевшие лошади. Факторы передачи - корм, вода, предметы ухода, кормушки и стены помещений, загрязнённые гноем и носовым истечением больных лошадей. Эпизоотии M. преим. наблюдают поздней осенью, зимой и ранней весной. Течение болезни чаще острое. Типичная форма M. характеризуется лихорадкой, угнетением, вялостью, уменьшением аппетита, увеличением подчелюстных лимфатич. узлов и образованием в них абсцессов. Диагноз ставят по эпизоотологич. данным, клинич. картине и результатам лабораторного исследования. Больных лошадей изолируют и лечат. Организуют строгое индивидуальное содержание, кормление и водопой животных. Помещения, предметы ухода за лошадьми, упряжь, кормушки тщательно дезинфицируют. Лит.: Б а к у л о в И. А , Мыт,
в кн.: Эпизоотология, под ред. P. Ф. Сосова. M., 1969.
МЫТИЩИ, город областного подчинения
в Московской обл. РСФСР. Ж.-д. станция в 19 км к С.-В. от Москвы,
от M.- ветки на Монино (30 км) и Пирогово (9 км). Нас. 125
тыс. чел. (1973; 60 тыс. чел. в 1939). Значит, пром. центр Подмосковья,
гл. отрасль - машиностроение. 3-ды: маш.-строит, (осн. в 1897; выпускает
автосамосвалы, вагоны для метро), приборостроит. и электромеханический.
Развита промышленность стройматериалов. Известностью пользуется з-д художеств,
литья, на котором были отлиты памятники Юрию Долгорукому, В. Маяковскому
и др. В M.- НИИ искусств, волокна и др. Моск. лесотех-нич. и кооперативный
ин-ты, маш.-строит. техникум, мед. уч-ще. Село Б. Мытищи возникло в 15
в., в кон. 18 в. от M. был проложен водопровод для снабжения Москвы водой
из мытищинских ключей. Город - с 1925.
МЫТНИК (Pedicularis), род растений сем. норичниковых. Многолетние, редко однолетние или двулетние полупаразитные травы с перисторассечёнными или перистораздельными листьями (нижние в розетке). Цветки неправильные, розово-фиолетовые, пурпуровые, жёлтые, белые, в колосовидных или кистевидных соцветиях; венчик двугубый. Плод - коробочка. Ок. 600 видов, в Сев. полушарии, преим. в Гималаях и Китае, 1 вид - в Юж. Америке. В СССР св. 110 видов, гл. обр. в Cp. Азии, по сухим каменистым склонам, альп. и субальпийским лугам. В сев. и средней полосе Европ. части широко распространён M. болотный (P. palust-ris), растёт на болотах, берегах водоёмов. Все его части ядовиты, содержат гликозид ауку-бин (ринантин); трава используется как инсектицид. Нек-рые виды M. разводят как декоративные. Мытник болотный.
МЫХЛИК Василий Ильич (р. 29.12.1922,
с. Солдатское, ныне Казанковского р-на Николаевской обл.), полковник (1953),
дважды Герой Сов. Союза (23.2.1945 и 29.6.1945). Чл. КПСС с 1944. В Советской
Армии с 1940. Окончил Вольское авиационно-технич. уч-ще им. Ленинского
комсомола (1941) и Воен.-возд. академию (1951). В Великую Отечеств, войну
1941-45-на Брянском, Зап., Ле-нингр., 3-м Белорус, фронтах. Был лётчиком,
командиром звена, зам. командира и командиром эскадрильи в частях 277-й
штурмовой авиадивизии. Произвёл 188 боевых вылетов. После войны - в ВВС.
С 1966 по болезни в запасе. Награждён орденом Ленина, 3 орденами Красного
Знамени, орденами Александра Невского, Отечеств, войны 1-й степени, Красной
Звезды и медалями. Бюст M. установлен в г. Кривой Рог.
МЫШЕВИДКИ (Phascogale), род сумчатых млекопитающих. Дл. тела до 22 см, хвоста - до 23 см. Окраска верха серая, на боках головы чёрный рисунок. 2 вида. Обитают в лесах Австралии. Ведут древесный образ жизни. Активны в сумерки и ночью. Питаются мелкими позвоночными и насекомыми. В Юж. Австралии распространена т а ф а (Ph. tapoatafa) - зверёк размером с белку, сходный по характеру питания с мелкими куньими (рис.). МЫШЕВИДНАЯ СОНЯ (Myomimus perso-natus),
млекопитающее семейства сонь отряда грызунов. В отличие от др. сонь,
внешне
похожа на мышь. Дл. тела , до 8 см, хвоста -7-8 см',
окраска
верха тела серая, низа - беловатая. Редкий вид; найден на Ю.-З. Туркмении,
в Иране и на Ю.-В. Болгарии. Обитает в предгорьях и на равнинах, на открытых
местах и в кустарниковых зарослях,
МЫШЕВИДНЫЕ ГРЫЗУНЫ, собирательное назв. мелких вредных грызунов сем. хомяков (Cricetidae) и мышей (Muridae) Типичные представители - лесные и серые полевки, степные пеструшки и хомячки, песчанки. Наносят большой вред нар. х-ву. Заселяют необрабатываемые земли и возделываемые поля. Прожорливы: суточный вес корма, потребляемого M. г., питающимися сочными частями растений, составляет 120- 300%, семенами - 30-100% от веса их тела. Очень плодовиты. Нек-рые виды могут размножаться круглый год. Самки серой полевки приносят за один помёт в среднем 5-7 (до 12) детёнышей и способны давать потомство через каждые 20-30 суток. Размножаются интенсивно. В жаркое время активны ночью и в сумерки, в холодное - днём. В течение вегетационного периода повреждают все с.-х. культуры, а особенно сильно зерновые и посевы многолетних трав. Зимой они выедают всходы озимых, объедают кору и корни деревьев в садах, питомниках, лесах, лесополосах, делают огромные запасы семян древесных культур. На пастбищах и сенокосах M. г. уничтожают ценные кормовые растения. Поселяясь в жилых постройках, на складах и в хранилищах, портят продукты, тару и сами постройки. Многие M. г.- переносчики возбудителей различных инфекционных и инвазионных заболеваний человека и домашних животных. Меры борьб ы-см. Дератизация. Лит.: Поляков И. Я, Вредные грызуны и борьба с ними, 2 изд., Л., 1968; Прохоров M. И, Микробиологический метод борьбы с вредными грызунами, Л , 1962. И. Я. Поляков. МЫШЕВИДНЫЙ ХОМЯЧОК (CaIo-myscus
bailwardi), млекопитающее семейства хомяков отр. грызунов. Дл. тела
до 8,5 см. Окраска песчано-жёлтая. В отличие от др. хомяков фауны
СССР, у M. х. хвост длиннее тела, с редкой кисточкой волос на конце, задние
ноги длиннее передних, защечных мешков нет. Обитает в горах юго-зап. Туркмении,
юж. Закавказья, а также Ирана.
МЫШЕХВОСТНИК (Myosurus), род однолетних травянистых растений сем. лютиковых. Листья узколинейные, в прикорневой розетке. Цветки мелкие, обоеполые, одиночные, на концах стеблей. Чашелистиков б. ч. 5; лепестки превращены в нектарники. Плод - многоорешек; плодики сидят на сильно вытянутом, почти цилиндрич. цветоложе. Ок. 6 (по др. данным, до 15) видов, преим. в умеренном поясе. В СССР 1 вид - M. маленький (M. minimus) - в Европ. части, на Кавказе, в Зап. Сибири и Cp. Азии; растёт как сорное растение в посевах, на залежах, у дорог, а также на сырых лугах и по берегам рек. МЫШЕЧНАЯ СИСТЕМА, мускульная система, совокупность сократимых элементов, мышечных клеток, объединенных обычно у животных и человека в мышцы и связанных между собой соединительной тканью. У одноклеточных, губок, кишечнополостных и нек-рых бескишечных ресничных червей M. с. нет. Движение их осуществляется за счет сокращения отростков эпителиальных клеток, колебаний жгутиков или ресничек. У большинства червей мышечные волокна обособлены от эпителия, хорошо развиты и вместе с наружными покровами образуют кожно-мускулъный мешок, состоящий обычно из наружных кольцевых и внутренних продольных волокон; кроме того, могут развиваться особые мышечные пучки у основания щетинок, в перегородках полости тела, вокруг кишечника, в стенках кровеносных сосудов. Как правило, у червей и моллюсков почти вся M. с. состоит из гладких мышц. У членистоногих развитая M. с. представлена сложной системой поперечно-полосатых мышц, прикрепленных к наружному скелету. У хордовых и особенно у позвоночных M. с. наиболее развита (в среднем 4/з- 4/2 массы тела). Она осуществляет осн. функции животных - от движения и поддерживания равновесия тела до транспортировки веществ (напр., пищи, крови) внутри организма. В M. с. происходит превращение химич. энергии как в механическую (см. Мышечное сокращение), так и тепловую, чем обеспечивается постоянный теплообмен с окружающей средой. M. с. хордовых делят на висцеральную мускулатуру, в осн. гладкую, происходящую из боковых пластинок и обслуживающую деятельность внутр. органов, и париетальную, или соматическую (скелетную), состоящую из поперечно-полосатых мышц, происходящую из мио-томов и реализующую взаимодействие организма с внеш. средой. В париетальной мускулатуре выделяют осевую, лежащую вдоль главной оси тела, и мускулатуру конечностей, происходящую из особых почек миотомов. Первично париетальная мускулатура служила для передвижения животного в воде при помощи горизонтальных изгибов тела и была представлена (как у совр. ланцетника) 2 продольными боковыми мышцами, разделёнными поперечными, изогнутыми соединительнотканными перегородками - миосептами - на отд. сегменты - миомеры, соответствующие мио-томам. С образованием у позвоночных головы и утратой гибкости передней части туловища передние миомеры редуцировались или частично приобрели иную функцию. Так, из 3 предушных головных миотомов у всех позвоночных развиваются 4 прямые и 2 косые мышцы, управляющие движениями глаза. У наземных позвоночных от них обособляется мышца, втягивающая глаз. Заушные головные миотомы сохраняются целиком лишь у круглоротых, образуя наджаберную и поджаберную мускулатуру. У рыб и наземных позвоночных передние заушные миотомы исчезают, наджаберная часть задних также исчезает или сильно редуцируется, а поджаберная хорошо развита и образует вместе с передними туловищными миомерами подъязычную мускулатуру, связывающую плечевой пояс с лежащими впереди висцеральными дугами (у рыб) или подъязычным аппаратом, а его - с подбородком (у наземных позвоночных); производное подъязычной мускулатуры - мышцы языка. Поверхностная мускулатура позвоночных животных. А - минога (круглоротое), Б - акула (рыба), В - протей (земноводное), Г - гаттерия (пресмыкающееся), Д - кошка (млекопитающее) 1 - жаберные щели; 2 - рот; 3 - наджаберная мускулатура; 4 - поджаберная мускулатура; 5 - туловищные миомеры; 6 - брызгальце; 7 - мышца, поднимающая нёбно-квадратный хрящ; 8 - мышца, приводящая нижнюю челюсть; 9 - общий сжиматель висцерального аппарата (дорзальный - сверху и вентральный - снизу); 10 - спинная мускулатура; // - брюшная мускулатура; 12 - мышца, поднимающая грудной плавник; 13 - опускающая его; 14 - поднимающая нижнюю челюсть; 15 - опускающая её; 16-нижнечелюстная мышца; 17 - трапециевидная; 18 - дельтовидная; 19 - KO-ракоидно-плечевая; 20 - двуглавая; 21 - разгибатели предплечья; 22 - трёхглавая; 23 - 25 - мышцы живота (23 - наружная косая; 24 - внутренняя косая; 25 - прямая); 26 - сжимающая шею; 27 - поперечно-остистые; 28 - длинная мышца спины; 29 - подвздош-но-крестцовые; 30 - грудная; 31 - височная; 32 - жевательная; 33 - Двубрюшная; 34 - грудино-сосцевидная; 35- надостистая; 36 - подостистая; 37 - поднимающая лопатку; 38 - трёхглавая плеча; 39 - нижние зубчатые; 40 - широкая мышца спины; 41 - портняжная; 42 - напрягающая фасцию бедра; 43 - средняя и большая ягодичные; 44 - полусухожильная; 45 - двуглавая бедра; 46 - икроножная. Собственно туловищная мускулатура кругл о р о т ы х представлена рядом мио-меров, состоящих из продольных волокон; поперечные миосепты выгнуты в форме ломаной линии. У р ы б появляется горизонтальная миосепта, отделяющая в каждом миомере более мощную спинную часть от брюшной. Спинная мускулатура сохраняет продольное направление волокон; в брюшной - оно сохраняется преим. по средней линии, а по бокам становится косым, что приводит к дифференциации различных слоев мышц. Изменение способа передвижения с переходом животных к наземной жизни и усиление конечностей привели к относит, уменьшению массы осевой мускулатуры, потере её строгой сегментации и приобретению ряда дополнит, функций. Спинная мускулатура в процессе эволюции изменяется сравнительно мало. У земноводных, как и у рыб, сохраняется парная сегментированная мышца, идущая от головы до конца хвоста. У пресмыкающихся она дифференцируется на 3 группы продольных мышц, изгибающих и фиксирующих позвоночник. Спереди выделяются мышцы, управляющие движениями головы и шеи, позади - мускулатура хвоста. У змей, по существу вернувшихся к исходному способу передвижения, спинная мускулатура хорошо развита и усложнена. У черепах и птиц в связи с ограничением подвижности спины она в значит, мере редуцируется. У млекопитающих сохраняются 3 группы спинных мышц, появившиеся у пресмыкающихся; кроме того, в поясничной области формируется мощная мышца, поддерживающая позвоночник в выпрямленном положении. Брюшная мускулатура наземных позвоночных служит гл. обр. для поддержания внутр. органов, сжатия полости тела, лёгочного дыхания и т. д., в связи с чем заметно усложнена. Под поперечными отростками и по бокам тел позвонков располагаются короткие подпозвоночные мышцы, служащие в области шеи и хвоста антагонистами спинной мускулатуры. По средней линии лежит парная прямая мышца живота, играющая особую роль у млекопитающих, как антагонист мощных разгибателей спины. В боковых стенках различают наружную и внутреннюю косые мышцы живота; наиболее глубоко лежит поперечная мышца живота. С развитием у амниот грудной клетки и аппарата рёберного дыхания в области рёбер дифференцируется ряд мышц, а у млекопитающих в связи с более интенсивным метаболизмом развивается ещё неск. групп мышц, способствующих дыханию,- верхняя зубчатая, лестничные и особенно диафрагма - производное шейных миото-мов. В передней части туловища от наружной косой мышцы живота обособляется у наземных позвоночных ещё ряд мышц, прикрепляющих плечевой пояс к осевому скелету (поскольку прямой костной связи между ними нет) и управляющих его движениями. Эта, т. н. вторичная, мускулатура передней конечности (в т. ч. и трапециевидная мышца висцерального происхождения) особенно развита у высших позвоночных. Наиболее изменчива в ходе эволюции позвоночных мускулатура конечностей. У рыб мускулатура парных плавников представлена лишь дорзальной, или спинной, мышцей, поднимающей плавник и отводящей его от средней линии (абдукция), и вентральной, брюшной,-· опускающей его, приводя к средней линии (аддукция). Отд. мышечные волокна могут выдвигать плавник вперёд (про-тракция) или оттягивать назад (ретракция). Непарные плавники управляются мускульными пучками, прикреплёнными к кожным лучам. Производными мускулатуры у рыб являются электрические органы. У наземных позвоночных конечности, превратившиеся в сложные рычаги, стали гл. орудиями передвижения и опоры. Поэтому их мускулатура дифференцировалась на многие мышцы, обеспечивающие сложные движения конечностей и удерживающие кости в фиксированном в суставах положении. Однако у зародышей можно проследить происхождение каждой из мышц из дорзальной или вентральной мускулатуры. У земноводных и примитивных наземных пресмыкающихся вентральная (брюшная) мускулатура особенно развита. Ещё более мощны эти мышцы в передней конечности птиц. В ходе эволюции млекопитающих конечности переместились вниз под тело, что резко изменило механику движения и опоры этих животных. Туловище в виде арки ложится на передние и задние конечности - подпорки. Изменился и способ ретракции задних конечностей - основа передвижения животного вперёд. Если у пресмыкающихся это достигается действием мышц вентрального происхождения, соединяющих хвост и бедро, то у млекопитающих, с изменением положения конечностей и потерей локомоторной функции хвоста, аналогичную функцию приняли на себя ягодичные мышцы - производные дорзальной мускулатуры, идущие от расширенных подвздошных костей таза к бедру. Приводящие мышцы (аддукторы) и др. заднебедренные мышцы - двуглавые, полусухожильные и полуперепончатые - становятся, в основном, ретракторами. У высших позвоночных поверхностные мышцы плечевого пояса дают начало подкожной мускулатуре туловища. Лит.: Шмальгаузен И. И., Основы сравнительной анатомии позвоночных животных, 4 изд., M., 1947; Беклемишев В. H., Основы сравнительной анатомии беспозвоночных, 3 изд., т. 2, M-, 1964. В. Б. Суханов.
МЫШЕЧНАЯ ТКАНЬ, ткань, составляющая осн. массу мышц и осуществляющая их сократит, функцию. Различают поперечнополосатую M. т. (скелетные и сердечная мышцы), гладкую и с двойной косой исчерченностью. Почти вся скелетная M. т. у позвоночных развивается из парных метамерных зачатков мускулатуры тела - миотомов. В процессе индивидуального развития организма одноядерные клетки - миобласты - интенсивно делятся и, сливаясь друг с другом, дают начало -"мышечным трубочкам", к-рые затем превращаются в дифференцированные мышечные волокна - осн. структурный элемент попереч-нополосатых мышц. Поперечная исчер-ченность волокон связана с чередованием в их многочисленных сократимых нитях - миофибриллах - участков с различными физико-химич. и оптич. свойствами. Гладкая M. т. кожи, стенок органов желудочно-кишечного тракта, мочеполовой системы и кровеносных сосудов развивается из клеток мезенхимы (слюнных, потовых и молочных желез - из эктодермы) и состоит из одноядерных ве-ретеновидных клеток (см. Гладкие мышцы). M. т. с двойной косой исчерченностью встречается относительно редко - у нек-рых червей и в запиратель-ных мышцах двустворчатых моллюсков. Исчерченность под острым углом (10- 15°) к продольной оси мышечного волокна возникает при совпадении (с нек-рым постоянным сдвигом) тёмных и светлых дисков соседних миофибрилл. Особенность этой M. т.- медленное сокращение и возможность длительного (многие часы) пребывания в сокращённом состоянии с небольшой затратой энергии. См. также ст. Мышцы и лит. при ней. МЫШЕЧНОЕ СОКРАЩЕНИЕ, укорочение мышцы, в результате к-рого она производит механич. работу. M. с. обеспечивает способность животных и человека к произвольным движениям. Наиболее важная составная часть мышечной ткани - белки (16,5-20,9%), в т. ч. контрактильные, обусловливающие способность мышцы к сокращению. Значит, интерес представляют механоактивные миофибриллярные белки, изучение к-рых было начато В. Кюне (1864). Важные данные, характеризующие физико-химич. и биохимич. свойства механоактивных мышечных белков, были получены А. Я. Данилевским (1881-88). В 1-й пол. 20 в. В. А. Энгельгардт и M. H. Любимова (1939) установили, что осн. контрактиль-ный белок мышц - миозин - обладает аденозинтрифосфатазной активностью, а А. Сент-Дьёрдьи и Ф. Б. Штрауб показали (1942-43), что входящий в состав миофибрилл белок состоит в основном из 2 компонентов - миозина и актина. Взаимодействие этих фибриллярных белков и лежит в основе феномена сокращения самых различных контрактильных органелл и органов движения (см. Мышцы). Периодич. изменение физич. состояния мышечных белков, обусловливающее возможность попеременного сокращения и расслабления мышц и выполнения ими механич. работы, очевидно, связано с определёнными дающими энергию биохимич. процессами. Энгельгардтом и Любимовой (1939-42) было обнаружено, что особым образом приготовленные из миозина нити при взаимодействии с раствором АТФ резко изменяют свои механич. свойства (эластичность и растяжимость). Одновременно происходит расщепление АТФ с образованием АДФ и неорганич. фосфата. Это открытие заложило фундамент для нового направления в биохимии- механохимии M. с. В дальнейшем Сент-Дьёрдьи и Штрауб показали, что истинным сократит, белком является не миозин, а его комплекс с актином - актомиозин. Сокращению при взаимодействии с АТФ подвергаются как вымоченные в воде или 50%-ном глицерине мышечные волокна, так и нити, приготовленные из актомиозиновых гелей (синерезис геля). Эти опыты подтверждают, что энергия, необходимая для сокращения мышц, освобождается в результате взаимодействия актомиозина с АТФ с расщеплением последнего на АДФ и Н3РО4. При этом освобождается большое количество энергии (8-10 ккал, или 33,5-41,9 кдж, на 1 моль АТФ). Однако истинный механизм этой реакции остаётся всё ещё неясным. Полагают, что терминальная фосфатная группировка АТФ при взаимодействии с актомиозином переносится на миозин без промежуточного образования тепла с образованием богатой энергией фосфорилированной формы актомиозина, способной к сокращению. Мол. масса миозина, определённая методом ультрацентрифугирования, близка к 500 тыс. Молекула миозина может быть расщеплена без разрыва кова-лентных связей на более мелкие субъединицы (рис. 1): 2 "тяжелые" полипептидные цепочки с мол. массой св. 210 000 и 2 (по др. данным, 3) коротких ("лёгких") полипептида с мол. массой ок. 20 000 каждый. Рис. 1. Структура молекулы миозина. Молекула состоит из 2 больших и 2 малых (коротких) цепей. Большие цепи образуют длинный "хвост" молекулы; ее "головка" состоит из концов больших цепей и 2 малых цепей. (Мол масса отдельных цепей определялась методом ультрацентрифугирования после дезагрегации молекулы миозина 5 M солянокислым гуанидином, а также трипсином.) По данным электронной микроскопии, молекула миозина состоит как бы из 2 частей - утолщённой "головки" и длинного " хвоста ". Общая длина молекулы - ок. 1600 А. Большое число соответственно расположенных в пространстве макромолекул миозина образует в поперечнопо-лосатом волокне толстые (миозиновые) нити. В образовании поперечных мостиков между толстыми (миозиновыми) и тонкими (актиновыми) нитями непосредственно участвуют, по-видимому, "головки" миозиновых молекул. Мол. масса мономера актина близка к 46 000 (ранее принималась ок. 70 000). Установлена и его первичная структура: число, природа и последовательность включения в полипептидную цепь аминокислотных остатков. Молекулы фибриллярного актина (Ф-актина) образованы 2 спиральными цепочками, состоящими из множества бусинок - молекул глобулярного актина (мономера актина, или Г-актина). В саркомерах поперечнополосатого волокна нити Ф-актина пространственно отграничены от нитей миозина. Взаимодействие систем двух типов нитей осуществляется за счёт энергии, освобождающейся при расщеплении АТФ в присутствии ионов Ca2+(рис. 2). T. к. при работе мышц АТФ постоянно потребляется, для длит, осуществления двухфазной мышечной деятельности необходимо непрерывное Рис. 2 Гипотетические схемы мышечного сокращения
CL
- конфигурация толстых и тонких нитей в расслабленной мышце; изменение
положения нитей при сокращении, 6 - модель скольжения (по X. Хаксли); восстановление АТФ - её ресинтез. Ресинтез АТФ из АДФ и H3PO4 сопряжен с рядом дающих энергию превращений. Важнейшие из них: 1) перенос фосфатной группы с фосфокреатина (КрФ) на АДФ. Эта реакция обеспечивает быстрый, происходящий уже во время сокращения мышцы, ресинтез АТФ за счёт потребления креатинфосфата; 2) глико-генолиз, или гликолиз (расщепление гликогена или глюкозы с образованием молочной к-ты); 3) тканевое дыхание (образование АТФ в митохондриях мышечных волокон за счет энергии окисления гл. обр. углеводов, жирных K-T и ненасыщенных фосфолипидов). Нек-рое количество АТФ может образовываться также в результате миокиназной реакции из АДФ: 2 АДФ <>АМФ + АТФ Фос-форилироваяие креатина за счет АТФ с образованием КрФ осуществляется в процессе гликолиза и тканевого дыхания. Ресинтез КрФ и гликогена происходит гл. обр. в фазе отдыха после расслабления мышцы. Скелетная мышца, находящаяся в анаэробных условиях или в условиях кислородного голодания (гипоксии), способна к выполнению некоторого кол-ва работы. Однако утомление в этих случаях наступает значительно раньше, чем в присутствии кислорода, и сопровождается накоплением в мышце молочной кислоты. А В. Палладиным, Д. Л. Фердманом, H. H. Яковлевым и др. получены данные о биохимической сущности тренировки мышц. С. E. Севериным продемонстрирована способность дипептидов (карнози-на, анзерина) восстанавливать работоспособность утомленных мышц и влиять на передачу нервных импульсов с нерва на мышцу. После сокращения, вызванного раздражением
с нерва или электрич. током, мышца вскоре переходит в расслабленное состояние,
хотя содержание АТФ в мышечных волокнах почти не меняется. Установлено,
что миофибриллы обладают способностью взаимодействовать с АТФ и сокращаться
в ее присутствии лишь при наличии в среде ионов Ca2+. Наибольшая
сократит, активность наблюдается при концентрации Ca2+ ок. 10~6
-10~5 моль. При понижении содержания Ca2+
до 10~7 моль или ниже мышечные волокна теряют способность
к укорочению и развитию напряжения (тянущей силы) в присутствии АТФ. По
совр. представлениям, в покоящейся мышце концентрация ионов Ca2+ поддерживается
ниже этой пороговой величины вследствие их связывания структурами (трубочками
и пузырьками) саркоплазматической сети. Связывание - это не простая адсорбция,
а активный физиологич. процесс, осуществляемый за счёт энергии, освобождающейся
при расщеплении АТФ в присутствии ионов Mg. Этот механизм получил название
Са-насоса (по аналогии с ^натриевым насосом"). T. о., пребывание
живой мышцы (при наличии в ней достаточного кол-ва АТФ) в расслабленном
состоянии - результат снижения под действием Са-насоса концентрации ионов
Ca2+ в среде, окружающей миофибриллы, ниже предела, при к-ром
ещё возможно проявление АТФ-азной активности и сократимости актомиозиновых
структур волокна. Сокращение волокна при раздражении с нерва (или электрич.
током) - результат внезапного изменения проницаемости и, как следствие,
выхода из цистерн И трубочек саркоплазматической сети и т. н. Т-системы
ионов Ca2+ в межфибриллярное пространство. Поперечные трубочки
Т-системы, расположенные на уровне Z-дисков и содержащие Ca2+,
сообщаются с поверхностной мембраной волокна; поэтому волна деполяризации
быстро распространяется по системе трубочек и достигает глубоко расположенных
участков волокна. После затухания нервного импульса в результате действия
Са-насоса концентрация Ca2+ в межфибриллярном пространстве быстро
снижается до пороговой величины и мышца переходит в исходное расслабленное
состояние, пока новый импульс не вызовет повторение всего цикла. Потерю
актомиозином способности расщеплять АТФ и сокращаться при снижении концентрации
ионов Ca2+ ниже 10-7 моль связывают с присутствием
в контрак-тильной системе особого белка - т Двухфазный механизм M. с. не исчерпывается изложенными представлениями. У нек-рых насекомых (жуки, пчелы, мухи, комары и др.) частота сокращений мышц крыльев много выше частоты поступающих нервных импульсов. Эти мышцы подчинены не нейрогенному, а миогенному ритму. Они могут совершать колебания (осциллировать) неск. сот раз в 1 сек. Осцилляция этих мышц не связана с изменением концентрации Ca2+ в саркоплазме мышечных волокон. Возможность автоматич. двухфазной деятельности клеточных органелл движения в присутствии АТФ можно наблюдать на клеточных моделях - сперматозоидах, мерцательном эпителии, унду-лирующих мембранах трипаносом и др. Осцилпяция органелл движения происходит с обычной для данного вида клеток скоростью при постоянной концентрации Ca и продолжается до тех пор, пока в растворе сохраняется известный избыток АТФ. Механизм такой осцилляции органелл движения, а также мио-фибрилл, по-видимому, может быть понят лишь исходя из существования взаимосвязи между ферментативной активностью (способностью расщеплять АТФ) и состоянием (конформаци-ей) макромолекул сократительного субстрата. Лит. см при ст. Мышцы. И. И. Иванов. МЫШЕЧНОЕ ЧУВСТВО, мышечно-суставная рецепция, проприорецепция, способность человека и животных воспринимать и оценивать изменение в относит, положении частей тела и их перемещение. На роль информации о положении той или иной части тела в пространстве и о степени сокращения каждой из мышц в регуляции движений и познании окружающей среды впервые указал И. M. Сеченов, назвавший M. ч. "тёмным мышечным чувством". Нервные импульсы, возникающие в мышечно-суставных (кинестетических) рецепторах - проприорецепто-рах (к ним относятся мышечные веретена, тельца Гольджи, а возможно, и Пачини) при сокращении и растяжении мышц, по чувствит. нервным волокнам достигают центр, нервной системы. Совокупность участвующих в анализе этой информации периферия, и центр, нервных образований наз. И. П. Павловым двигательным анализатором. Совершенство и тонкость координации двигательных реакций, в т. ч. и локомоций, осуществляемых животным и человеком, объясняются накоплением в течение жизни организма всё новых связей между нейронами двигательного анализатора и др. анализаторов (зрительного, слухового и др.). M. ч. играет важнейшую роль в развитии восприятий организма, т. к. служит осн. контролем остальных органов чувств. Так, зрительная оценка удалённости к.-л. предмета вырабатывается с помощью M. ч. при приближении к предмету. О. M. Бенюмов. MЫШИ (Muridae), семейство млекопитающих отр. грызунов. Дл. тела 5-50 см; хвоста до 45 см. Подразделяется на 2 подсемейства - мышиные (Murinae) и авст-рал. водяные крысы (Hydromyinae); нек-рые зоологи выделяют 5 и более подсемейств. Всего 80 совр. родов и 12 вымерших с более 400 видов; распространены всесветно, большинство видов- в лесах тропиков и субтропиков. Завезены в Сев. и Юж. Америку и на многие острова. В СССР 11 видов из 5 родов. Активны в сумерках и ночью. Большинство ведёт полуназемный образ жизни, питаясь семенами, часть - животной пищей. Нек-рые - синантропные животные (домовая мышь, нек-рые крысы). Размножаются в тёплое время года; в жилищах человека - круглогодично. Половозрелыми становятся в 1,5-3 месяца. M.- природные носители большого числа паразитов и хранители возбудителей многих болезней человека и домашних животных, включая опасные инфекции. Вредят зерновому и лесному х-ву, повреждают материалы и продукты питания. Наибольший вред приносят M. из родов MUS и Apodemus (напр., домовая мышь) и крысы. Меры борьбы: механические, химические и бактериологические. Лит.: Аргиропуло А. И., Сем. Muridae - Мыши, М.- Л., 1940 (Фауна СССР, т. 3, в. 5); M i s о nn е X., African and indo-australian Muridae, evolutionary trends, "Annales du Musee Royal de 1'Afri-que Centrale", 1969, ser. 8, № 172. И. М. Громов. МЫШИНЫЙ ГИАЦИНТ, род растений сем. лилейных; то же, что гадючий лук. МЫШИНЫЙ ГОРОШЕК, вика многоцветковая (Vicia cracca), многолетнее травянистое растение семейства бобовых из рода горошек. Стебель дл. до 1,5л, ветвистый. Листья парноперисто-сложные, с 5-20 парами линейных или продолговато-яйцевидных листочков , на верхушке с усиком, цепляющимся за окружающие травы. Цветки от сине-фиолетовых до голубых, редко белые, в многоцветковых кистевидных соцветиях. Мышиный горошек; а - цветок, 6 - плоды. И. H. Мышкин. Плод - боб с 4-8 семенами. Произрастает в Евразии, Сев. Африке и как заносное в Сев. Америке; в СССР - почти по всей территории, но гл. обр. в лесной и лесостепной зонах. Растёт по лугам, кустарникам, опушкам, разреженным лесам, у жилья, иногда как сорное в посевах зерновых культур. Ценная кормовая трава, хорошо поедается скотом на пастбищах и в сене. Медонос. Горьковатый вкус семян M. г. обусловлен присутствием в них гликозида вицианина. Лит.: Кормовые растения сенокосов и пастбищ СССР, т. 2, М.- Л., 1951. МЫШКИН Ипполит Никитич [22.1(3.2). 1848, Псков,- 26.1(7.2). 1885, Шлиссельбург], русский революционер, народник. Сын воен. писаря, унтер-офицера, и крепостной крестьянки. Окончил школу кантонистов в Пскове (1860) и училище колонновожатых (1864) в Петербурге. В 1864-68 в чине унтер-офицера служил топографом в Петербурге и правительств, стенографом в Москве. Оставив воен. службу, сдал экзамен на звание домашнего учителя, работал стенографом, сотрудничал в периодич. печати. В 1874) приобрёл в Москве типографию, в к-рой в 1874 совместно с П. И. Войнаралъским организовал печатание запрещённых на-роднич. изданий. После разгрома типографии M. скрылся за границу. Весной 1875 отправился в Сибирь, намереваясь освободить H. Г. Чернышевского. В Ви-люйске после вооруж. сопротивления арестован в июле 1875, отправлен в Петербург и заключён в Петропавловскую крепость. M.- один из главных обвиняемых по "процессу 193-х". 15 нояб. 1877 выступил на суде с яркой революц. речью. Приговорён к 10 годам каторжных работ. Наказание отбывал в центр, каторжных тюрьмах (Новобелгородской и Ново-борисоглебской), в 1881 отправлен в Вост. Сибирь. В нач. 1882 приговорён дополнительно к 15 годам каторжных работ за речь, произнесённую в Иркутской тюрьме при прощании с умершим Л. А. Дмоховским. Бежал с Карийской каторги 19 апр. 1882, арестован во Владивостоке 24 мая, заключён в Шлиссель-бургскую крепость. Расстрелян по приговору воен. суда за протест против тюремного режима. Лит.: Ленин В. И·, Что делать?,
Поли, собр. соч., 5 изд., т. 5; Революционное народничество 70-х гг. XIX
в., т. 1, 1870- 1875, M., 1964; Язвицкий В., Непобеждённый пленник, M.,
1972; Остро-в е МЫШКИН Николай Филиппович (1864-1948), советский вет. акушёр, проф. (1919). Окончил Юрьевский (Дерпт-ский) вет. ин-т (1891). Организовал (1919) и возглавил в Московском вет. ин-те первую в СССР кафедру акушерства. Разработал эффективный метод клинич. диагностики ранних сроков стельности коров, получивший распространение в вет. практике СССР. Соч.: Акушерство и гинекология сельскохозяйственных животных, 4 изд., M., 1943; Наружные методы определения беременности сельскохозяйственных животных, M., 1936. Лит.: Коропов В. M, История ветеринарии в СССР, M., 1954. МЫШКИНО, посёлок гор. типа, центр
Мышкинского р-на Ярославской обл. РСФСР. Пристань на лев. берегу р. Волги,
в 21 км к Ю. от ж.-д. ст. Волга (на линии Сонково - Рыбинск) и в
126 км к С.-З. от г. Ярославля. Кирпичный завод, птицефабрика. В
районе M.- компрессорная станция газопровода "Сияние Севера".
МЫШЛАЕВСКИЙ Александр Захарьевич [12(24).3.1856 - 1920], русский воен. деятель и историк, ген. от инфантерии (1912). Окончил Михайловское арт. уч-ще (1877) и Академию Генштаба (1884). С 1898 проф. Академии Генштаба и в 1899-1904 одновременно нач. Воен.-учёного архива Гл. штаба. В 1908-09 нач. Гл. штаба, в марте - сент. 1909 - нач. Ген. штаба. С 1913 пом. наместника на Кавказе по воен. части. В начале 1-й мировой войны 1914-18 пом. главнокомандующего Кавказской армией, с марта 1915 в отставке. С дек. 1915 пред. Металлургич. к-та и уполномоченный по делам металлургич. пром-сти Особого совещания по обороне гос-ва. С авг. 1916 гл. уполномоченный по снабжению металлами. В марте - июне 1917 командующий войсками Казанского воен. округа. В окт. 1917 прикомандирован к штабу Кавказского фронта, с дек. 1917 пред. Кавказской воен.-историч. комиссии. Занимался изучением рус. воен. истории 18 в., отстаивая идею самобытности рус. воен. иск-ва. M. принадлежит большая заслуга в публикации значит, числа архивных материалов. Соч.: Северная война на Ингерманланд-ском и Финляндском театрах в 1708-1714, СПБ, 1893; Петр Великий. Военные законы и инструкции, СПБ, 1894; Война с Турцией 1711 г., СПБ, 1898; Офицерский вопрос в XVIII веке, СПБ, 1899; Две катастрофы. Суворов в Швейцарии, Петр на Пруте, СПБ, 1901; Северная война 1708 г. От реки Уллы и Березины за рекой Днепр, СПБ, 1901. Л Б. Леонидов
МЫШЛЕНИЕ, процесс отражения объективной действительности, составляющий высшую ступень человеческого познания. Хотя M. имеет своим единств, источником ощущения, оно переходит границы непосредственно-чувственного отражения и позволяет получать знание о таких объектах, свойствах и отношениях реального мира, к-рые не могут быть непосредственно восприняты человеком (см. Ф. Энгельс, в кн.: Маркс К. и Энгельс Ф., Соч., 2 изд., т. 20, с. 554- 555). M. составляет предмет изучения теории познания и логики, психологии и нейрофизиологии; оно изучается также в кибернетике в связи с задачами тех-нич. моделирования мыслительных операций. M. является функцией мозга и в этом смысле представляет собой естеств. процесс. Однако каждый отд. человек становится субъектом M., лишь овладевая языком, понятиями, логикой, к-рые суть продукты развития обществ, практики. Даже задачи, к-рые он ставит перед своим M., порождаются обществ, условиями его жизни. T. о., M. человека имеет общественно-историч. природу. Вопрос о природе M., взаимоотношении M. (сознания) и бытия был центральной проблемой на протяжении всей истории филос. мысли (см. Основной вопрос философии, Материализм, Идеализм). Конкретно-науч. изучение M., развернувшееся в 19 в., находилось под влиянием представлений формальной логики и учения об ассоциациях. Психологич. анализ M. сводился гл. обр. к выделению отд. мыслит, процессов: сравнения, абстрагирования признаков, классификации. В духе формальной "школьной" логики освещался вопрос о природе понятий, к-рые изображались как продукт наслаивания друг на друга чувственных образов и представлений, выделения в них общих признаков и устранения несовпадающих. Соответственно процессы M. представлялись в виде сложных сочетаний представлений и понятий. Это понимание M. наталкивалось на неразрешимые трудности при объяснении, напр., целенаправленного и творч. характера мыслит, процессов. Натуралистич. и ме-ханистич. представления о M. развивались позднее бихевиоризмом, к-рый истолковывал мыслит, деятельность как совокупность беззвучных речевых навыков, формирующихся по схеме "стимул- реакция". Этим ассоцианистским и ме-ханистич. концепциям M. противостояли идеалистич. направления, подчёркивавшие несводимость мыслит, процессов к ассоциированию отд. представлений, свойственную им безобразность и подчинённость их "детерминирующим тенденциям" (Вюрцбургская школа). С др. стороны, представители теории целостных форм (гешталътпсихология) понимали M. как процесс "переструктурирования" субъектом проблемной ситуации, открывающий новые заключённые в ней отношения и функциональные связи и невыводимый из опыта поведения и накопленных в нём ассоциаций. Истолковывая M. как "автохтонный" самопорождающийся процесс, представители гештальтпсихологии смыкались при этом с интуитивизмом. Общей чертой этих концепций является их антиисторизм, отказ от изучения происхождения и историч. развития человеческого M. Только в нач. 20 в. появились работы, в к-рых систематизировались накопленные данные о M. народов, стоящих на относительно низких ступенях общественно-экономич. и культурного развития. Эти работы показали несостоятельность положения о неизменности законов M. и внесли идею о качественных изменениях M. в процессе его историч. развития (Л. Леви-Брюлъ, Франция, и др.). Одновременно начались экспериментальные исследования зачатков M. в животном мире (В. Кёлер, Германия; P. Йеркс, США; H. H. Ладыгина-Коте, СССР), выявившие у высших животных процессы поведения, сходные с человеческим мышлением ("практический интеллект" или, по И. П. Павлову, "ручное мышление"). Эти исследования не только углубили понимание генетич. корней M., но и дали толчок изучению M., протекающего в форме внеш. действия с объектами ("наглядно-действенное М.", "технический интеллект") и у человека. Открытие мыслит, деятельности в виде внеш. действий в сложных предметных ситуациях, операций с наглядными схемами, макетами конструкций и т. д. разрушало старое представление о M. как только внутреннем, словесноло-гич. процессе и вело к признанию существования у человека различных форм высокоразвитого M., теснейшим образом переплетающихся между собой и переходящих друг в друга. Науч. диалектико-материалистич. понимание природы M. было дано в трудах классиков марксизма. Отвергая взгляды на M. как на проявление особого духовного начала, марксизм преодолевает вместе с тем ограниченность метафизич. материализма, его созерцательность и сведение мыслит, деятельности к элементарным процессам анализа и обобщения чувственных впечатлений. Рассматривая M. как продукт общественно-историч. развития, как особую форму человеческой деятельности, марксизм подчёркивает изначальную связь M. с практич. деятельностью людей. "Производство идей, представлений, сознания первоначально непосредственно вплетено в материальную деятельность... Образование представлений, мышление, духовное общение людей является здесь ещё непосредственным порождением материального общения людей" (M а р к с К. и Энгельс Ф., там же, т. 3, с. 24). Труд посредством орудий ставит человека не только перед материальными объектами, но и перед их взаимодействием, в процессе к-рого обнаруживаются свойства, недоступные непосредственно нашим ощущениям и постигаемые лишь опосредствованно, путём умозаключений. Познават. результаты предметных действий закрепляются в словесной форме и, передаваясь в процессе речевого общения др. людям, входят в систему знаний, составляющих содержание сознания коллектива, общества. Языковая форма выражения создаёт условие, благодаря к-рому отд. звенья внешнепредметной познават. деятельности могут выполняться уже только во внутр. речевом плане, в плане сознания. Исходные чувственные данные и практич. действие опосредствуются всё более длинным рядом мыслит, процессов, приобретающих впоследствии способность отделяться от внеш. практич. деятельности. При этом обществ, разделение труда, развитие частной собственности и дифференциация общества на антагонистич. классы приводят к отрыву умственного труда от физического, так что внутр. мыслит, деятельность начинает всё более противопоставляться деятельности материальной. Это противопоставление закрепляется позднее в идеалистич. теориях M. Утрата M. в его развитых формах прямой и непосредств. связи с практич. деятельностью приводит к тому, что оно может давать ложные, иллюзорные знания; это ставит проблему критерия истинности M., адекватности его результатов объективной реальности. Таким критерием является практика: теоретич. результаты, к к-рым приходит M., должны проверяться в практич. деятельности и в эксперименте. Однако под практикой следует понимать здесь не индивидуальную, а совокупную обществ, практику, что делает необходимым подчинение процессов M. определ. правилам или предписаниям, выработанным в историч. опыте познания. Созданная человечеством система такого рода правил и предписаний ("законов" M.) и составляет содержание особой дисциплины - логики. В противоположность идеалистич. взглядам на логич. законы как имманентно присущие M. марксизм рассматривает их как обобщённое отражение объективных отношений действительности, осваиваемых практикой. "...Практическая деятельность человека миллиарды раз должна была приводить сознание человека к повторению разных логических фигур, дабы эти фигуры м о г-л и получить значение аксиом" (Ленин В. И., Поли. собр. соч., 5 изд., т. 29, с. 172). Обществ, практика служит не только критерием истинности M., но является также той основой, на к-рой вырастают логич. правила и законы. Поэтому M. не может быть сведено к совокупности мыслит, операций, образующих его состав, и, следовательно, к "мышлению" логич. машин, выполняющих лишь те процессы, к-рые так или иначе задаются им человеком. Машины являются лишь "...созданными человеческой рукой органами человеческого мозга..." (M арке К., см. Маркс К. и Энгельс Ф., Соч., 2 изд., т. 46, ч. 2, с. 215), истинным же субъектом M. остаётся управляющий им человек. Огромное усложнение задач, решаемых совр. наукой, потребовало дальнейшего развития логич. аппарата M., что привело к созданию новых направлений формальной логики. Изучение M., однако, не может ограничиваться только исследованием формально-логич. правил. Оно включает в себя прежде всего проблемы отношения M. к объективной реальности и общего метода познания. Единство познават. и логич. аспектов M. нашло своё наиболее полное выражение в марксистской диалектической логике, представляющей собой учение о развитии, самодвижении предмета познания, как оно отражается в движении понятий M. A. H. Леонтьев. Расстройства мышления находят своё отражение в речи, письме и изобразит, творчестве. Возможны заметные колебания темпа M. Ускорение потока мыслей на фоне эмоционального возбуждения способно достичь степени "скачки идей"- нецеленаправленных, поверхностных. Заторможенность M., напротив, характеризуется скудным и замедленным возникновением представлений, образованием понятий и суждений, что согласуется с подавленным настроением больного и субъективным ощущением интеллектуального опустошения. Другие проявления нарушений M.: его бессвязность (инкогеренция); инертность с преобладанием к.-л. одной мысли, одного представления (персеверация); тугоподвиж-ность, обстоятельность, при которых изложение существа тонет в массе ненужных подробностей, а также бесплодное, далёкое от реальной действительности M.; вычурность понятий, тенденция к символике, парадоксальное применение общеизвестных понятий и неоправданное словотворчество; параллельное течение неск. мыслей; обрывы мыслей и, наконец, полная разорванность содержания M., но с грамматически правильной структурой предложений. В дифференциальной диагностике психич. заболеваний симптомы расстройств M. имеют большое значение (см., напр., Шизофрения). Б. И. Франкштейн. Лит.: Маркс К. и Энгельс Ф., Немецкая
идеология, Соч., 2 изд., т. 3; Энгельс Ф., Диалектика природы, там же,
т. 20; Ленин В. И., Философские тетради, Поли. собр. соч., 5 изд., т. 29;
Б и н э А., Механизм мышления, пер. с англ., Од., 1894; Психология
мышления, СПБ, 1914 (Новые идеи в философии, в. 16); ЛевН-Брюль Л., Первобытное
мышлеч ние, пер, с франц., M., 1930; Валлон А., От действия к мысли, пер.
с франц., M., 1956; Выготский Л. С., Мышление и речь. Избранные психологические
исследования, M., 1956; Рубинштейн С. Л., О мышлении и путях его исследования,
M., 1958; Пономарёв Я. А., Психология творческого мышления, M., 1960; 3
е й-гарник Б. В, Патология мышления, JM.], 1962; Психология мышления, сб.
пер., M., 1965; Тихомиров О. К., Структура мыслительной деятельности человека,
M., 1969; Пиаже Ж , Психология интеллекта, в его KH : Избранные психологические
труды, [пер. с франц.], M , 1969; Леонтьев A H., Проблемы развития психики,
3 изд., M., 1972; Ac h N., Uber die Begriffsbildung, Bamberg, 1921; К о
f f-k а К., Bemerkungen zur Denkpsychologie, "Psychologische Forschung",
1927, Bd 9, [H. 1,2]; Hartley D. M., Observation on man, his frame, his
duty and his expectations, 6 ed , L., 1934; A study of thinking, N. Y ,
1956.
МЫШOBКИ (Sicista), род млекопитающих сем. тушканчиков отр. грызунов. Внешне похожи на мышь. Дл. тела до 8 см', голый хвост длиннее тела. 10 видов; в СССР - 6. Населяют равнинные и горные леса и открытые ландшафты умеренного и субтропического поясов Европы и Азии. Активны в сумерках; питаются насекомыми и семенами. Приносят 1 помёт в году. Зимой впадают в спячку. Наиболее известны: лесная M. (S. betulina) и степная M. (S. sub-tilis). Лесная мышовка.
МЫШЦЫ, мускулатура скелетная и внутренних органов (висцеральная), обеспечивающая у животных и человека выполнение ряда важнейших физиологич. функций: перемещение тела или отдельных его частей в пространстве, кровообращение, дыхание, передвижение пищ. кашицы в пищеварит. органах, поддержание тонуса сосудов, выделение экскрементов и т. д. Сократит, функция всех типов M. обусловлена превращением в мышечных волокнах химич. энергии определённых биохимич. процессов в механич. работу. Однако сокращение скелетных M. и мускулатуры внутренних органов - лишь частный случай более общей закономерности - механо-химич. активности живых структур. По-видимому, в основе самых различных проявлений этой активности - сокращения хвоста сперматозоида, движения ресничек инфузорий, расхождения хромосом во время митоза, впрыскивания в бактерию фаговой ДНК и т. д.- лежит один и тот же молекулярный механизм, связанный с возможностью изменения конформации или взаимного расположения фибриллярных структур контрактильных белков. Типы мышц. Морфологи различают 2 осн. типа M.: поперечнополосатые мышцы и гладкие мышцы. К первым относится вся скелетная мускулатура позвоночных животных и человека (см. илл. на вклейке), обеспечивающая возможность выполнения произвольных движений, M. языка, верх, трети пищевода и некоторые др., M. сердца (миокард), имеющая свои особенности (состав белков, характер сокращения и др.), а также M. членистоногих и нек-рых др. бес-лозвонрчных. К гладким M. принадлежит большая часть мускулатуры беспозвоночных животных и мышечные слои внутр. органов и стенок кровеносных сосудов позвоночных животных и человека, обеспечивающие возможность выполнения ряда важнейших физиологич. функций. Нек-рые гистологи, изучающие M. беспозвоночных, выделяют и 3-й тип M.- с двойной косой исчерченностью (см. Мышечная ткань). Структурными элемент а-м и всех типов M. являются мышечные волокна (рис. 1). Поперечнополосатые мышечные волокна в скелетных M. образуют пучки, соединённые друг с другом прослойками соединительной ткани. Рис. 1. Поперечнополосатые мышечные волокна человека: а - отрезки двух волокон; б - поперечный разрез волокна, миофибрил-лы образуют скопления, разграниченные саркоплазмой; в - поперечный разрез волокна с равномерным распределением миофибрилл. Своими концами мышечные волокна сплетаются с сухожильными волокнами, через посредство к-рых мышечная тяга передаётся на кости скелета. Волокна поперечнополосатых M. представляют собой гигантские многоядерные клетки, диаметр к-рых варьирует от 10 до 100мкм, а длина часто соответствует длине M., достигая, напр., в нек-рых M. человека 12 см. Волокно покрыто эластичной оболочкой - сарколеммой и состоит из саркоплазмы, структурными элементами к-рой являются такие органоиды, как митохондрии, рибосомы, трубочки и пузырьки саркоплазматической сети и т. н. Т-системы (рис. 2), различные включения и т. д. Рис. 2. Электронная микрофотография саркоплазматической сети и Т-системы мышечного волокна. В саркоплазме обычно в форме пучков расположено множество нитевидных образований толщиной от 0,5 до неск. мкм - миофибрилл, обладающих, как и всё волокно в целом, поперечной исчерченностью. Каждая миофибрилла разделена на неск. сот участков длиной 2,5-3 мкм, наз. саркомерами. Каждый саркомер, в свою очередь, состоит из чередующихся участков - дисков, обладающих неодинаковой оптической плотностью и придающих миофибриллам и мышечному волокну в целом характерную поперечную исчерченность, чётко обнаруживаемую при наблюдении в фазовоконтрастном микроскопе. Более тёмные диски обладают способностью к двойному лучепреломлению и наз. анизотропными, или дисками Л. Более светлые диски не обладают этой способностью и наз. изотропными, или дисками /. Среднюю часть диска А занимает зона более слабого двойного лучепреломления - зона H. Диск / делится на 2 равные части тёмной Z-пластинкой, отграничивающей один саркомер от другого. В каждом сарко-мере имеется два типа нитей (филамен-тов), состоящих из мышечных белков: толстые миозиновые и тонкие - акти- Рис. 3. Строение сар-комера поперечно-полосатого мышечного волокна А - электронная микрофотография (малое увеличение), на которой чётко видна структура саркоме-ра: Б - схема сар-комера; В - электронная микрофотография с высокой разрешающей способностью; Г - поперечное сечение саркомера на различных уровнях, видно положение толстых и тонких нитей в различных участках покоящегося саркомера (по X. Хаксли). Скелетные мышцы человека. 1. Вид спереди: 1 - затылочно лобная; 2 - круговая мышца рта; 3 -подбородочная; 4 - грудино-подъязычная; S - трапециевидная; 6 - трёхглавая мышца плеча; 7 - прямая мышца живота; 8 -наружная косая мышца живота; 9 - подвздошно-поясничная; 10 - мышца, натягивающая широкую фасцию; 11 - гребешковая; 12 - длинная приводящая; 13 - портняжная; 14 - тонкая; 15 - прямая мышца бедра; 16 - медиальная широкая мышца бедра; 17 - боковая широкая мышца бедра; 18 - отводящая большой палец стопы; 19 - длинный разгибатель пальцев (сухожилие); 20 - длинный разгибатель пальцев; 21 - передняя большеберцовая; 22 - камбаловидная; 23 - икроножная; 24 - короткий разгибатель большого пальца кисти; 25 - длинный разгибатель большого пальца кисти; 26 - локтевой сгибатель запястья; 27 - короткий лучевой разгибатель запястья; 28 - разгибатель пальцев; 29 - лучевой сгибатель запястья; 30 - длинный лучевой разгибатель запястья; 31 - плече-лучевая; 32 - трёхглавая мышца плеча; 33 - двуглавая мышца плеча; 34 - передняя зубчатая; 35 - большая грудная; 36 - дельтовидная; 37 - средняя лестничная; 38 - передняя лестничная; 39 и 40 - грудино-ключично-сосцевидная; 41 - опускающая угол рта; 42 - жевательная; 43 - большая скуловая; 44 - височная. 2. Вид сзади: 1 - затылочно-лобная; 2 - трапециевидная; 3 - дельтовидная; 4 - трёхглавая мышца плеча; 5 - двуглавая мышца плеча; 6 - круглый пронатор; 7 - длинная ладонная; 8 - лучевой сгибатель запястья; 9 - поверхностный сгибатель пальцев; 10 - плече-лучевая; 11-локтевой сгибатель запястья; 12 - короткая мышца, отводящая большой палец кисти; 13 и 14 - полуперепончатая; 15 - полусухожильная; 16 - тонкая; 17 - двуглавая мышца бедра; 18 - полуперепончатая; 19 - икроножная; 20 - камбаловидная; 21 - большая ягодичная; 22 - натягивающая широкую фасцию; 23 - средняя ягодичная; 24 - наружная косая живота; 25 - широчайшая мышца спины; 26 - зубчатая передняя; 27 - большая круглая; 28 - подгребешковая; 29 - малая круглая; 30 - плече-лучевая; 31 - грудино-ключично-сосцевидная; 32 - ремённая мышца головы; 33 - жевательная; 34-полуостистая; 35 - височная. К ст. Мышцы.
К ст. Насекомоядные растения. Ia и 16. Жирянка обыкновенная (Pinguicula vulgaris). 2. Росянка капская (Drosera capensis). 3. Росянка круглолистная (Drosera rotundifolia). 4. Росолист (Drosophyllum lusitanicum). 5. Саррацения (Sarracenia drummondii). 6. Венерина мухоловка (Dionea muscipula). 7. Непентес (Nepenthes destillatoria). 8. Пузырчатка обыкновенная (Utricularia vulgaris). 9. Альдрованда (Aldrovanda vesiculosa). новые (рис. 3). Неск. иную структуру имеют
гладкие мышечные волокна. Они представляют собой веретенообразные одноядерные
клетки, лишённые поперечной исчерченности. Длина их обычно достигает 50-250
мкм
(в
матке - до 500 мкм), ширина - 4-8 мкм', мио-филаменты в них
обычно не объединены в обособленные миофибриллы, а расположены по длине
волокна в виде множества одиночных актиновых нитей (рис. 4). Упорядоченная
система миозиновых нитей в гладкомышечных клетках отсутствует. В гладкой
мускулатуре моллюсков наиболее важную роль в осуществлении запирательной
функции играют, по-видимому, парамиозиновые волокна (тропомиозин А).
Рис. 4 Электронная микрофотография гладкомышечного
волокна (увеличено в 60 000 раз). Видны актиновые нити (показаны стрелками).
Химический состав мышц колеблется в зависимости
от вида и возраста животного, типа и функционального состояния M. и ряда
др. факторов. Осн. вещества, входящие в состав по-перечнополосатых M. человека
и животных, и их содержание (в % к сырой массе) представлены ниже:
В среднем ок. 75% сырой массы M. составляет
вода. Осн. количество плотных веществ приходится на долю белков. Различают
белки миофибриллярные (сократительные) - миозин, актин и их комплекс
- актомиозин, тропомиозин и ряд т. н. минорных белков (а и Функциональные особенности поперечнополосатых мышц. Поперечнополосатые M. богато снабжены разл. нервами, с помощью к-рых осуществляется регуляция мышечной деятельности со стороны нервных центров. Важнейшие из них: двигательные нервы, проводящие к M. импульсы, вызывающие её возбуждение и сокращение; чувствительные нервы, по к-рым от M. к нервным центрам поступает информация о её состоянии, и, наконец, адаптационно-трофические волокна симпатической нервной системы, воздействующие на обмен веществ и замедляющие развитие утомления M. (см. Адаптационна-трофическая функция). Каждая веточка двигательного нерва, иннервирующего целую группу мышечных волокон, образующих т. н. моторную единицу, доходит до отдельного мышечного волокна. Все мышечные волокна, входящие в состав такой единицы, сокращаются при возбуждении практически одновременно. Под влиянием нервного импульса в окончаниях двигательного нерва высвобождается медиатор-ацетилхолин, взаимодействующий с холинорецептором пост-синапгической мембраны (см. Синапсы). В результате этого происходит повышение проницаемости мембраны для ионов Na и К, что, в свою очередь, обусловливает её деполяризацию (появление постсинап-тического потенциала). После этого на соседних участках мембраны мышечного волокна возникает волна возбуждения (волна электроотрицательности), к-рая распространяется по скелетному мышечному волокну обычно со скоростью неск. м в 1 сек. В результате возбуждения M. изменяет свои эластические свойства. Если точки прикрепления M. не фиксированы неподвижно, происходит её укорочение (сокращение). При этом M. производит определённую механич. работу. Если точки прикрепления M. неподвижны, в ней развивается напряжение. Между возникновением возбуждения и появлением волны сокращения или волны напряжения протекает нек-рое время, наз. латентным периодом. Сокращение M. сопровождается выделением тепла, к-рое продолжается в течение определённого времени и после их расслабления. В M. млекопитающих и человека установлено существование "медленных" мышечных волокон (к ним принадлежат "красные", содержащие дыхат. пигмент миоглобин) и "быстрых" ("белых", не имеющих миоглобина), различающихся скоростью проведения волны сокращения и её продолжительностью. В "медленных" волокнах млекопитающих длительность волны сокращения примерно в 5 раз больше, а скорость проведения в 2 раза меньше, чем в "быстрых" волокнах. Почти все скелетные M. относятся к смешанному типу, т. е. содержат как "быстрые", так и "медленные" волокна. В зависимости от характера раздражения возникает либо одиночное - фазное - сокращение мышечных волокон, либо длительное - тетаническое. Тетанус возникает в случае поступления в M. серии раздражений с такой частотой, при к-рой каждое последующее раздражение ещё застаёт M. в состоянии сокращения, вследствие чего происходит суммирование сократит, волн. H. E. Введенский установил, что увеличение частоты раздражений вызывает возрастание тетануса, но лишь до известного предела, наз. им "оптимумом". Дальнейшее учащение раздражений уменьшает тетаническое сокращение (пессимум). Развитие тетануса имеет большое значение при сокращении "медленных" мышечных волокон. В M. с преобладанием "быстрых" волокон макс, сокращение - обычно результат суммации сокращений всех моторных единиц, в к-рые нервные импульсы поступают, как правило, неодновременно, асинхронно. В поперечнополосатых M. установлено также существование т. н. чисто тонических волокон, к-рые особенно широко представлены в M. земноводных и пресмыкающихся. Тонические волокна участвуют в поддержании "неутомляемого" мышечного тонуса. Тоническим сокращением наз. медленно развивающееся слитное сокращение, способное длительно поддерживаться без значит, энергетич. затрат и выражающееся в "неутомляемом" противодействии внешним силам, стремящимся растянуть мышечный орган. Тонические волокна реагируют на нервный импульс волной сокращения лишь локально (в месте раздражения). Тем не менее, благодаря большому числу концевых двигательные бляшек тоническое волокно может возбуждаться и сокращаться всё целиком. Сокращение таких волокон развивается настолько медленно, что уже при весьма малых частотах раздражения отдельные волны сокращения накладываются друг на друга и сливаются в длительно поддерживающееся укорочение. Длительное противодействие тонических волокон, а также медленных фазных волокон растягивающим усилиям обеспечивается не только упругим напряжением, но и возрастанием вязкости мышечных белков. Для характеристики сократительной функции M. пользуются понятием "абсолютной силы", к-рая является величиной, пропорциональной сечению M., направленной перпендикулярно её волокнам, и выражается в кг/см2. Так, напр., абс. сила двуглавой M. человека равна 11,4, икроножной - 5,9 кг/см2. Систематическая усиленная работа M. (тренировка) увеличивает их массу, силу и работоспособность. Однако чрезмерная работа приводит к развитию утомления, т. е. к падению работоспособности M. Бездеятельность M. ведёт к их атрофии. Функциональные особенности гладких мышц. Гладкие M. внутренних органов по характеру иннервации, возбуждения и сокращения существенно отличаются от скелетных M. Волны возбуждения и сокращения протекают в гладких M. в очень замедленном темпе. Развитие состояния "неутомляемого" тонуса гладких M. связано, как и в тонических скелетных волокнах, с замедленностью сократительных волн, сливающихся друг с другом даже при редких ритмических раздражениях. Для гладких M. характерна также способность к автоматизму, т. е. к деятельности, не связанной с поступлением в 'M. нервных импульсов из центральной нервной системы. Установлено, что способностью к ритмическому самопроизвольному возбуждению и сокращению обладают не только нервные клетки, имеющиеся в гладкий M., но и сами гладко-мышечные клетки Своеобразие сскратительной функции гладких M. позвоночных животных определяется не только особенностями их иннервации и гистологич. строения, но и спецификой их химич. состава: более низким содержанием контрактильных белков (актомиозина), макроэргических соединений, в частности АТФ, низкой АТФ-азной активностью миозина, наличием в них водорастворимой модификации актомиозина - тоноактомиозина и т. д. Существенное значение для организма имеет способность гладких мышц изменять длину без повышения напряжения (наполнение полых органов, например мочевого пузыря, желудка и др.). И. И. Иванов. Скелетные мышцы человека, различные по форме, величине, положению, составляют св. 40% массы его тела. При сокращении происходит укорочение M., к-рое может достигать 60% их длины; чем длиннее M. (самая длинная M. тела - портняжная - достигает 50 см), тем больше размах движений. Сокращение куполообразной M. (напр., диафрагмы) обусловливает её уплощение, сокращение кольцеобразных M. (сфинктеров) сопровождается сужением или закрытием отверстия. M. радиального направления, наоборот, вызывают при сокращении расширение отверстий. Если M. расположены между костными выступами и кожей, их сокращение обусловливает изменение кожного рельефа. Все скелетные, или соматические (от греч. soma - тело), M. по топо-графо-анатомич. принципу могут быть разделены на M. головы, среди к-рых различают мимические и жевательные M., воздействующие на нижнюю челюсть, M. шеи, туловища и конечностей. M. туловища покрывают грудную клетку, составляют стенки брюшной полости, вследствие чего их делят на M. груди, живота и спины. Расчленённость скелета конечностей служит основанием для выделения соответствующих групп M.: для верхней конечности - это M. плечевого пояса, плеча, предплечья и кисти; для нижней конечности - M. тазового пояса, бедра, голени, стопы. У человека ок. 500 M., связанных со скелетом. Среди них одни крупные (напр., четырёхглавая M. бедра), другие - мелкие (напр., короткие мышцы спины). Совместная работа M. выполняется по принципу синергизма, хотя отд. функциональные группы M. при выполнении определ. движений работают как антагонисты. Так, спереди на плече находятся двуглавая и плечевая M., выполняющие сгибание предплечья в локтевом суставе, а сзади располагается трёхглавая M. плеча, сокращение к-рой вызывает противоположное движение - разгибание предплечья. В суставах шаровидной формы происходят простые и сложные движения. Напр., в тазобедренном суставе сгибание бедра вызывает пояснично-подвздошная M., разгибание - большая ягодичная. Бедро отводится при сокращении средней и малой ягодичных M., а приводится с помощью пяти M. медиальной группы бедра. По окружности тазобедренного сустава локализуются также M., к-рые обусловливают вращение бедра внутрь и кнаружи. Наиболее мощные M. размещаются на туловище. Это M. спины - выпрямитель туловища, M. живота, составляющие у человека особую формацию - брюшной пресс. В связи с вертикальным положением тела M. нижней конечности человека стали более сильными, поскольку, кроме участия в локомоции, они обеспечивают опору тела. M. верхней конечности в процессе эволюции, напротив, сделались более ловкими, гарантирующими выполнение быстрых и точных движений. На основе анализа пространственного положения и функциональной деятельности M. совр. наука пользуется также следующим их объединением: группа M., осуществляющая движения туловища, головы и шеи; группа M., осуществляющая движения плечевого пояса и свободной верх, конечности; M. нижней конечности. В пределах этих групп выделяются более мелкие ансамбли. В. В. Куприянов. Патология мыщц. Нарушения сократит, функции M. и их способности к развитию и поддержанию тонуса наблюдаются при гипертонии, инфаркте миокарда, мио-дистрофии, атонии матки, кишечника, мочевого пузыря, при разл. формах параличей (напр., после перенесённого полиомиелита) и др. Патология, изменения функций мышечных органов могут возникать в связи с нарушениями нервной или гуморальной регуляции, повреждениями отд. M. или их участков (напр., при инфаркте миокарда) и, наконец, на клеточном и субклеточном уровнях. При этом может иметь место нарушение обмена веществ (прежде всего ферментной системы регенерации макроэргических соединений - гл. обр. АТФ) или изменение белкового сократит, субстрата. Указанные изменения могут быть обусловлены недостаточным образованием мышечных белков на почве нарушения синтеза соответствующих информационных, или матричных, РНК, т. е. врождённых дефектов в структуре ДНК хромосомного аппарата клеток. Последняя группа заболеваний, т. о., относится к числу наследственных заболеваний. Саркоплазматич. белки скелетных и гладких M. представляют интерес не только с точки зрения возможного участия их в развитии вязкого последействия. Многие из них обладают ферментативной активностью и участвуют в клеточном метаболизме. При повреждении мышечных органов, напр, при инфаркте миокарда или нарушении проницаемости поверхностных мембран мышечных волокон, ферменты (креатинкиназа, лактат-дегидрогеназа, альдолаза, аминотрансфе-разы и др.) могут выходить в кровь. T. о., определение активности этих ферментов в плазме крови при ряде заболеваний (инфаркт миокарда, миопатии и др.) представляет серьёзный клинич. интерес. Лит.: Энгельгардт В. А., Ферментативные
и механические свойства белков мышц, -"Успехи современной биологии", 1941,
т. 14, в. 2; С е н т - Д ж и о р д ь и А., О мышечной деятельности, пер.
с англ., M., 1947; Иванов И. И., Юрьев В. А., Биохимия и патобиохимия мышц,
Л., 1961; Поглазов Б. Ф., Структура и функции сократительных белков, M.,
1965; X а и а-ш и Т., Как клетки движутся, в кн.: Живая клетка, пер. с
англ., 2 изд., M., 1966; Хаксли Г., Механизм мышечного сокращения, в сб.:
Молекулы и клетки, пер. с англ., в. 2, M., 1967; Смит Д., Летательные мышцы
насекомых, там же; Бендолл Дж., Мышцы, молекулы и движение, пер. с англ.,
M., 1970; Арронет H. И., Мышечные и клеточные сократительные (двигательные)
модели, Л., 1971; Л ё в и А., С и к е виц Ф., Структура и функции клетки,
пер. с англ., M., 1971; Иванов И- И., Некоторые актуальные проблемы эволюционной
биохимии мышц, "Журнал эволюционной биохимии и физиологии", 1972, т. 8,
№ 3; Gibbons I. R., The biochemistry of motility, "Annual Review of Biochemistry",
1968, v. 37, p. 521. И. И. Иванов.
|
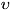 течения
жидкости или газа к скорости звука а в данной точке потока M
=
v/a. Обычно М-ч. наз. Маха числом. В сов. литературе М-ч.
иногда наз. числом Маиев-ского. ,
течения
жидкости или газа к скорости звука а в данной точке потока M
=
v/a. Обычно М-ч. наз. Маха числом. В сов. литературе М-ч.
иногда наз. числом Маиев-ского. ,
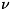 .
5, N. Y.- L.- Toronto, 1959.
.
5, N. Y.- L.- Toronto, 1959.
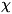 o-зяйственные,
туалетные и технические M.; они бывают твёрдыми, мягкими, жидкими и порошкообразными.
Жировым сырьём в произ-ве M. служат
жиры животные и жирные масла
растительные, а также жирозаменители - синтетич. жирные к-ты, канифоль,
нафтеновые кислоты, талловое масло.
Твёрдые сорта M. получают из твёрдых
жиров и саломасов - отверждённых гидрогенизацией растит, масел или жидких
жиров морских животных (см. Жиров гидрогенизация). Сырьём для жидких
M. служат в основном жидкие растит, масла, наряду с к-рыми используют жирозаменители.
В произ-ве туалетного
мыла жидкого жирозаменители не применяют.
o-зяйственные,
туалетные и технические M.; они бывают твёрдыми, мягкими, жидкими и порошкообразными.
Жировым сырьём в произ-ве M. служат
жиры животные и жирные масла
растительные, а также жирозаменители - синтетич. жирные к-ты, канифоль,
нафтеновые кислоты, талловое масло.
Твёрдые сорта M. получают из твёрдых
жиров и саломасов - отверждённых гидрогенизацией растит, масел или жидких
жиров морских животных (см. Жиров гидрогенизация). Сырьём для жидких
M. служат в основном жидкие растит, масла, наряду с к-рыми используют жирозаменители.
В произ-ве туалетного
мыла жидкого жирозаменители не применяют.
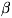 - модель скручивания (по Подольскому).
- модель скручивания (по Подольскому).
 oпон
и н а. При его отсутствии актомио-зин реагирует in vitro (в пробирке) с
АТФ практически и в отсутствии Ca2+. В физиологических условиях
(in vivo) тропонин - постоянный компонент кон-трактильной системы мышечного
волокна (тропонин-тропомиозиновый комплекс).
oпон
и н а. При его отсутствии актомио-зин реагирует in vitro (в пробирке) с
АТФ практически и в отсутствии Ca2+. В физиологических условиях
(in vivo) тропонин - постоянный компонент кон-трактильной системы мышечного
волокна (тропонин-тропомиозиновый комплекс).