БОЛЬШАЯ СОВЕТСКАЯ ЭНЦИКЛОПЕДИЯ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ
|
БОЛЬШАЯ СОВЕТСКАЯ ЭНЦИКЛОПЕДИЯ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ |
ГЕНЕТИКА-ГЕНУЭЗСКАЯ
"ГЕНЕТИКА", научный журнал АН СССР. Издаётся в Москве с 1965. В журнале помещаются статьи, обзоры, сообщения по вопросам молекулярной, химич. и радиационной генетики, мутационной теории, генетики популяций, ци-тогенетики, взаимоотношений генотипа и среды, теории селекции, частной генетики и селекции животных, растений и микроорганизмов, экспериментальной полиплоидии, гетерозиса, отбора и гибридизации, генетики человека. К статьям даётся резюме на англ, языке. Тираж (1971) ок. 3,2 тыс. экз. Журнал переводится и издаётся на англ, языке в США. ГЕНЕТИКА ЖИВОТНЫХ, раздел генетики,
изучающий наследственность и изменчивость преим. с.-х., а также домашних
и диких животных. Основывается на общегенетич. принципах и положениях и
использует в основном такие методы общей генетики, как гибридологический,
цитологический, популяционный, онтогенетический, математико-статистический,
близнецовый и др.
Чаще всего у животных наблюдается независимое наследование
признаков, обусловленное большим числом хромосом. Напр., диплоидное число
хромосом у уток 80, у собак и кур по 78, лошадей 66, кр. рог. скота и коз
по 60, овец 54, кроликов 44, свиней 40, лисиц 38, норок 30. Осн. методом
изучения наследования признаков служит гибридологический анализ. Этот метод
позволил выяснить характер наследования мн. морфологич., физиол. и биохим.
особенностей, часто зависящих только от одной или неск. пар генов.
Большое внимание уделяется генетике биохим. свойств молока,
крови животных, в частности иммуногенетике, результаты к-рой используются
для контроля за родословными племенных животных, уточнения их происхождения
в спорных случаях и т. д. Установлена возможность с помощью изучения генов,
обусловливающих биохим. свойства, вести анализ структуры пород, их линий
и отродий, судить о степени однотипности пород и т. п. Продолжаются исследования
коррелятивных связей этих генов с продуктивностью, плодовитостью и жизнеспособностью
животных.
Генетич. объяснение получили встречающиеся у животных
морфологич. недостатки и недоразвитие отдельных органов. Известно, что
мн. из пороков развития (бульдоговидность, карликовость и водянка головы
у телят, безногость у поросят, безволосость у телят и крольчат и др.) определяются
т. н. летальными и полулетальными генами. Особи - носители таких генов,
или гибнут, или обладают низкой жизнеспособностью. Появление животных с
такими недостатками объясняется тем, что в стадах встречаются особи, внешне
нормальные и вполне жизнеспособные, но гетерозиготные по генам, определяющим
эти недостатки. При скрещивании таких гетерозиготных особей друг с другом
в потомстве появляются нежизнеспособные формы, гомозиготные по летальным
или полулетальным генам.
Летальное или полулетальное действие могут оказывать и
гены, обусловливающие полезные в хоз. отношении признаки. Классич. пример
этого - доминантный ген, определяющий у каракульских ягнят серую окраску
- ши-рази, к-рый одновременно оказывается рецессивным в отношении жизнеспособности
особей.
Новым и перспективным направлением Г. ж. является генетика
устойчивости к нек-рым инфекционным, инвазионным и грибковым заболеваниям.
Известны генетически обусловленные различия устойчивости животных к маститу,
туберкулёзу, ящуру, пироплазмозу и др.
Развитие у животных количественных признаков - скороспелости,
величины удоя, содержания жира в молоке, настрига шерсти, яйценоскости
и др. - зависит от деятельности мн. систем организма. Этим объясняется
сложная генетич. природа этих признаков. Установлено, что количественные
признаки определяются совокупным действием мн. генов с однозначным действием.
Последние могут различаться по степени доминирования, вплоть до сверхдоминантных
генов, вызывающих гетерозис в первом поколении помесей. Для изучения количественных
признаков пользуются математико-статистич. методами.
Породы и внутрипородные группы с.-х. животных (линии,
семейства и т. д.) - всегда популяции, в к-рых происходит расщепление по
мн. генам. Популяц. метод позволяет изучить распространение отдельных генов
в популяциях животных. В простейших случаях, при расщеплении в популяции
по одному или немногим генам, параметрами, характеризующими популяции,
служат частоты отдельных генов. При анализе признаков, зависящих от мн.
генов, частоты отдельных генов не могут быть установлены, и тогда пользуются
коэффициентом наследуемости - отношением генотипич. изменчивости количественного
признака к его общей фе-нотипич. изменчивости. Значения коэфф. наследуемости
(от 0 до 1) зависят от специфики признаков, для к-рых они устанавливаются,
а также от степени вырав-ненности условий содержания и кормления и от методов
разведения животных. Значение коэфф. наследуемости позволяет найти наиболее
подходящие методы селекции и прогнозировать их результаты.
ГЕНЕТИКА МЕДИЦИНСКАЯ, раздел генетики человека, изучающий наследственные заболевания и методы их предупреждения, диагностики и лечения. Существование заболеваний, передающихся по наследству (гемофилия и др.), а также тот факт, что браки между родственниками увеличивают частоту появления в потомстве наследственных заболеваний, были известны давно. В нач. 20 в. исследовалась гл. обр. соотносительная роль наследственности и среды в происхождении не только нормальных, но и патологич. признаков человека. В России Г. м. зародилась лишь при Советской власти и значит, развития достигла в 30-е гг. В Медико-генетическом ин-те, возглавлявшемся С. Г. Левитом, успешно изучалось наследование сахарного диабета, язвенной и гипертонич. болезней и др. С. Н. Давиденков и его школа всесторонне исследовали наследств, болезни нервной системы. При изучении наследств, болезней Г. м. пользуется всеми методами генетики человека: генеалогическим (заключается в составлении родословной, что позволяет выявить соотношение между здоровыми и больными членами семьи пробанда, т. е. больного, для к-рого составляется родословная), близнецовым (см. Близнецы), цитологическим, биохимическим, иммунологическим. В 60-х гг. 20 в. большое значение приобрели цитогенетич. методы исследования (хромосом и полового хроматина) и биохим. тесты. Выяснилось, что наследств, болезни зависят от изменений в хромосомах половых клеток,- это могут быть изменения структуры генов, хромосомные перестройки (делеции, транслокации, дупликации и др.) или количественные изменения в хромосомном наборе (добавление или потеря одной или неск. хромосом). Так, нарушением числа или структуры хромосом - половых или аутосом - обусловлены синдром Тернера - Шерешевского, Дауна болезнь, синдромы Клайнфельтера и кошачьего крика, трисомии 18 (Эдвардса) и D (Патау) и др. При др. наследств, заболеваниях обнаружить видимые дефекты в хромосомном наборе не удаётся. В таких случаях, по-видимому, происходят генные мутации или неблагоприятное сочетание различных генов. Проявление нек-рых наследств, болезней зависит от средовых факторов, к-рые могут способствовать развитию наследств, предрасположения или полностью его подавить. В задачу Г. м. входит выявление соответств. условий среды (в т. ч. леч. средств, диеты и др.). Успехи в развитии Г. м. сделали возможными предупреждение и лечение ряда наследств, болезней. Один из эффективных методов такого предупреждения - медико-г енетическое консультирование с предсказанием риска появления больного в потомстве лиц, страдающих данным заболеванием или имеющих больного родственника. Достижения биохим. генетики раскрыли первичные (молекулярные) дефекты при мн. наследственно обусловленных аномалиях обмена веществ, что способствовало развитию методов экспресс-диагностики, позволяющих быстро и рано выявлять больных и лечить мн. прежде неизлечимые наследств, болезни. Так, напр., подбором спец. диеты возможно предупредить развитие фенил-кетонурии (фенилпировиноградной олигофрении) и нек-рых др. наследств, болезней. В иных случаях лечение состоит во введении в организм извне веществ, не образующихся в нём в результате генетич. дефекта. Мн. генетич. дефекты исправляются своеврем. хирургич. вмешательством или педагогич. коррекцией. Во мн. странах организованы клиники и научные ин-ты, занимающиеся изучением наследственной патологии человека; в СССР - Ин-т мед. генетики АМН СССР. Ежегодно публикуется ок. 3 тыс. работ по Г. м. Вопросы Г. м. освещают журналы Acta geneticae medicae et gemello-logiae (Roma, с 1952); American Journal of Human Genetics (Baltimore, с 1949); Annals of Human Genetics (L., с 1954); Journal de genetique humaine (Geneve, с 1952); Journal of Medical Genetics (L., с 1964); Excerpta medica, Section. 22, Human genetics (Amst., с 1962). В СССР статьи по Г. м. публикуются во мн. мед. и биол. журналах (напр., в журн. Генетика, Цитология, Цитология и генетика). Е.
Ф. Давиденкова.
ГЕНЕТИКА МИКРООРГАНИЗМОВ, раздел общей генетики, в к-ром объектом исследования служат бактерии, микроскопии, грибы, актинофаги, вирусы животных и растений, бактериофаги и др. микроорганизмы. До 40-х гг. 20 в. считалось, что, поскольку у микроорганизмов нет ядерного аппарата и мейоза, на них не распространяются Менделя законы и хромосомная теория наследственности. С начала 40-х гг. микроорганизмы становятся объектом интенсивных геиетич. исследований. Именно на них были решены мн. кардинальные вопросы совр. генетики. Так, первое указание на то, что материальным носителем наследственности служит дезоксирибонуклеиновая кислота (ДНК), было получено в опытах на пневмококках (амер. генетики О. Т. Эйвери, К. Мак-Леод и М. Маккарти). Примерно в то же время были начаты интенсивные генетич. исследования на хлебной плесени - нейроспоре. Изучение многочисл. биохим. мутантов нейроспоры (Дж. У. Бидл и Э. Л. Тей-тем, США) привело к установлению очень важного положения: один ген - один фермент (ныне это положение более точно формулируется так: один ген - одна полипептидная цепь). Генетич. исследования микроорганизмов особенно интенсивно стали развиваться после того, как амер. генетики С. Лурия и М. Дельбрюк показали на кишечной палочке (Escherichia coli), что и бактерии подчиняются мутационным закономерностям (см. Изменчивость, Мутации). Ранее существовавшее представление об адекватной, адаптивной изменчивости у бактерий возникло вследствие методич. ошибки, заключавшейся в изучении культуры как единицы изменчивости. Был предложен новый принцип изучения изменчивости у бактерий - клональный анализ, т. е. изучение потомства одной клетки - родоначальницы клона. Важной вехой в развитии Г. м. явился разработанный амер. генетиками Дж. и Э. Ледербергами метод реплик, или отпечатков, позволивший доказать, что мутации возникают у бактерий независимо от условий культивирования, и, кроме того, значительно упростивший приёмы отбора вариантов микроорганизмов с желаемыми свойствами. Оказалось, что в больших популяциях бактериальных клеток мутации возникают спонтанно. В 1946 был открыт половой процесс у бактерий (конъюгация), что позволило применить для их исследования генетический анализ. В рсзультате установлены наличие у бактерий рекомбинации, существование у них генетич. групп сцепления и построены гепетич. карты их хромосом. Почти одновременно был открыт парасексуальный процесс у грибов (Г. Понтекорво, Великобритания), что расширило возможности генетич. анализа грибов, не имеющих полового цикла размножения. Вскоре в генетич. исследования были вовлечены бактериофаги и др. вирусы (в частности, вирус табачной мозаики - ВТМ). Был открыт эффект переноса генетич. информации от одной бактериальной клетки к другой при посредстве бактериофага - генетич. трансдукция, что положило начало изучению генетических взаимоотношений в системе фаг - бактерия (Дж. Ледерберг, Н. Зиндер, США). Вслед за тем была обнаружена рекомбинация у фагов (А. Херши и М. Дельбрюк, США). Если использование бактерий в качестве объекта генетич. исследований резко повысило разрешающую способность генетич. анализа, то благодаря фагам удалось перейти к изучению явлений наследственности на молекулярном уровне. Большое значение имели исследования ВТМ (нем. генетики Г. Шустер и А. Гирер), позволившие вызвать генетич. эффект в опытах с чистой рибо-нуклеиновой к-той (РНК), к-рая сохраняла ннфекционность и при нанесении на листья табака вызывала в клетках образование полноценных частиц ВТМ. Исходя из общих принципов исследования генетпч. процессов у микроорганизмов, для каждой группы разработаны спец. методы изучения с учётом их особенностей . Генетич. механизмы у грибов и водорослей, сохранивших половой процесс, имеют ряд особенностей. Главная из них состоит в том, что продукты мейоза (споры) остаются соединёнными в определ. порядке, и после раздельного высева этих спор можно непосредственно изучать генотип каждого продукта мейоза. Этот метод, называемый тетрадным анализом, дополняет статистич. методы изучения процесса расщепления. Применение генетнч. анализа к организмам, у к-рых отсутствует половой процесс, стало возможным после открытия у них парасексуальных процессов, отличающихся большим разнообразием. Так, у несовершенных грибов при срастании гиф, принадлежащих двум генетически различным штаммам, происходит объединение и затем слияние двух гаплоидных ядер в одно днплопдное; в этой системе изредка возможен обмен генетич. материалом. Особенность полового процесса у бактерии состоит в том, что в клетку-реципиент передаётся, как правило, только часть генетич. материала из клетки-донора, в результате чего образуется частично диплоидная зигота (т. н. мерозигота). У бактерий известно неск. механизмов передачи генетнч. материала. Наиболее совершенная форма полового процесса у бактерий - конъюгация, детально изученная у кишечной палочки. Конъюгация происходит при непосредств. контакте между двумя клетками, если в одной из них присутствует специфич. половой фактор, или фактор скрещиваемости (фертильности, плодовитости). Половой фактор (см. Эписомы) содержит ДНК и может существовать в клетке либо в автономном, либо в интегрированном состоянии (включённым в геном клетки). В первом случае при конъюгации в клетку-реципиент переходит только половой фактор. Во втором случае половой фактор способствует направленному переносу генетич. материала из клетки-донора в клетку-реципиент. Как правило, при этом происходит передача только части генома донора и лишь крайне редко передается вся хромосома донора вместе с включённым в неё половым фактором. Между фрагментом донорной ДНК и ДНК реципиента может произойти обмен гомологичными генетич. участками- кроссинговер, приводящий к возникновению рекомбинантов, т. е. клеток с изменённым сочетанием признаков. Генетич. анализ рекомбинантов кишечной палочки позволил установить у неё существование одной группы сцепления, определить линейное расположение большого числа генов в её хромосоме и построить кольцевую генетич. карту (см. Генетические карты хромосом). Перенос генетич. материала при конъюгации - строго ориентированный процесс, при к-ром последовательность передачи генов (а значит, и вероятность их участия в кроссннговере) целиком зависит от расположения генов в хромосоме и точки интеграции (включения) полового фактора. При переходе полового фактора в автономное состояние гены, расположенные на хромосоме рядом с точкой интеграции, могут объединиться с половым фактором н в дальнейшем передаваться с ним как единое целое, превращая клетки-реципиенты в диплоиды по данному генетич. участку. Этот процесс переноса генов совместно с половым фактором, паз. сексдукцией, также может привести к возникновению рекомбинантов. Др. механизм возникновения рекомбинантов у бактерий - трансдукция - осуществляется при посредстве т. н. умеренных бактериофагов, к-рые способны к особому виду симбиоза с бактериями - лизогении. В лизогенных бактериях ДНК умеренного фага интегрирована с ДНК бактериальной клетки и ре-плицируется одновременно с ней. Такая скрытая форма присутствия фага (профаг) может сохраняться в течение мн. клеточных поколений, однако изредка профаг переходит в вегетативное состояние (т. е. начинает размножаться) и разрушает бактерию. При этом возможны захват небольшого фрагмента ДНК клетки-хозяина и последующий его перенос в др. клетку, в к-рой перенесённый участок генома может вступить в генетич. обмен с гомологичной областью клетки-реципиента. Обычно при трансдукцпи передаются гены, расположенные в непосредств. близости от места локализации профага в хромосоме бактерии. Однако нек-рые фаги осуществляют трансдукцию, при к-рой любой участок генома бактерии с равной вероятностью может быть перенесён в др. клетку. Иногда сам процесс лизогенизацни, т. е. включения умеренного фага в геном бактерии, может сопровождаться приобретением клеткой новых свойств (см. Лизогенная конверсия), напр, вирулентности. Ещё один тип полового процесса у бактерий, называемый трансформацией,- перенос генетич. материала без посредства полового фактора или умеренного бактериофага с последующим возникновением рекомбинантов (вследствие генетич. обмена между пооникшим в клетку фрагментом ДНК и ДНК клетки-реципиента). Особенности генетических механизмов у вирусов бактерий - бактериофагов - делают их весьма удобной моделью для изучения воспроизведения и функционирования генетич. материала. Они очень просто устроены, быстро размножаются и имеют очень короткий жизненный цикл; поэтому генетика бактериофагов, в особенности фагов Т2, Т 4 и /\., исследована весьма детально. Бактериофаги скрещивают, заражая бактерии смесью двух или неск. мутантов фага. В этом случае, кроме исходных фаговых частиц, появляются рекомбинанты с изменёнными сочетаниями признаков. С помощью рекомбинационного анализа удалось построить генетич. карты для ряда бактериофагов. Оказалось, что молекула ДНК фага является его хромосомой. Изучение тонкой структуры гена, проведённое на фаге Т4 (С. Бензер, США), показало существование большого числа участков внутри гена, способных изменяться (мутировать) с разной частотой под действием различных мутагенов. Генетика вирусов животных и растений в значит, мере основывается на успехах в области генетики бактериофагов, но из-за технич. трудностей ещё не получила достаточного развития. Возможность получения рекомбинантов была показана у ДНК-содержащих вирусов группы оспы - осповакцины (при смешанном заражении клеток различными представителями этой группы), у вируса герпеса (между различными вариантами этого вируса), а также между обезьяньим опухолеродным вирусом SV40 и различными представителями адено-вирусов. У РНК-содержащих вирусов животных показана возможность получения рекомбинантов между мутантами вируса ящура и полиомиелита, а также между различными вариантами вируса гриппа. Последнее открытие имеет особое значение, т. к. показывает возможные пути изменчивости этого вируса в природе. Из вирусов растений лучше всего изучен вирус табачной мозаики (ВТМ). В частности, полностью расшифрована последовательность аминокислот в белке ВТМ; удалось установить характер аминокислотных замещений, возникающих в белках оболочки у разных мутантов ВТМ. Работы, выполненные на ВТМ, явились важным этапом в изучении как механизма мутагенеза, так и природы генетического кода. В связи с развитием новой отрасли народного хозяйства - микробиологической промышленности - возникла прикладная Г. м., наз. также селекцией микроорганизмов. В сферу исследований были вовлечены новые формы микроорганизмов: пенициллы (Penicillium chrysogenum), актиномицеты (Actinomyces streptomycini, Act. rimosus и др.), актинофаги. У пенициллов и аспергиллов открыт парасексуальный процесс, у актиномицетов изучен механизм рекомбинации, открыты генетич. рекомбинация у актинофагов, генетич. трансдукция у актиномицетов. Проведены обширные исследования индуцированной изменчивости количеств, признаков у актиномицетов. В Сов. Союзе в работе по селекции микроорганизмов применяются такие генетич. методы, как получение индуцированных мутаций, гибридизация и заражение актиномицетов актинофагами. В итоге выведены высокоактивные штаммы, позволившие во много раз увеличить производство антибиотиков, аминокислот, витаминов и др. биологически активных веществ. В связи с возрастающим значением Г. м. и необходимостью развития микробиол. пром-сти в 1968 в Москве был организован Всесоюзный н.-и. ин-т генетики и селекции пром. микроорганизмов Главмикробиопрома, ставший ведущим науч. центром в этой области. Проблемы генетики и селекции микроорганизмов разрабатываются и в др. науч. учреждениях Москвы (Ин-т эпидемиологии и микробиологии им. Н. Ф. Гамалеи АМН СССР, Ин-т общей генетики и Ин-т атомной энергии им. И. В. Курчатова АН СССР), Ленинграда (Физико-технич. ин-т им. А. Ф. Иоффе АН СССР, кафедра генетики ЛГУ), Киева, Еревана и др. Г.м. сыграла важную роль в развитии совр. генетики, дополнив ряд положений генетики высших организмов. Г. м., в свою очередь, стала основой для развития молекулярной генетики. Лит.: X э и с У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Гольд-Фарб Д. М., Введение в генетику бактерий, М., 1966; Захаров И. А. и Квитко К. В., Генетика микроорганизмов, Л., 1967; Алиханян С. И., Современная генетика, М., 1967; его же, Селекция промышленных микроорганизмов, М., 1968; Браун В., Генетика бактерий, пер. с англ., М., 1968; Генетические основы селекции микроорганизмов, М., 1969. С. И.
Алиханян, Л. Н. Майсурян.
ГЕНЕТИКА ПОВЕДЕНИЯ, область науки о поведении, основывающаяся на законах генетики и изучающая, в какой степени и каким образом различия в поведении определяются наследственными факторами. Осн. методы исследования Г. п. на экспериментальных животных - селекция в сочетании с инбридингом (близкородственное скрещивание), при помощи к-рых изучаются механизмы наследования форм поведения, на человеке - статистич. и генеалогич. анализ в сочетании с близнецовым и цитогенетич. методами. Зависимость поведения от наследств, факторов - генное управление и контроль поведения - исследуется на различных уровнях организации живого: в биоценозах, популяциях, сообществах, на уровне организма, а также на физиологическом (орган, ткань, клетка) и молекулярном уровнях. Исследования Г. п. имеют существ, значение для учения об индивидуальных различиях высшей нервной деятельности и выявления относит, роли врождённых и индивидуально приобретённых особенностей поведения, для объяснения роли генетически обусловленных особенностей поведения животных в популяции (для обществ, животных - в стаде, стае и т. п.), а также для создания экспериментальных моделей нервных болезней. Лит.: Крушинский Л. В., Генетика и феногенетнка поведения животных, в кн.: Актуальные вопросы современной генетики, М., 1966; Гэйто Д ж., Молекулярная психобиологпя, пер. с англ., М., 1969; Fа1сопег D. S., Introduction to quantitative genetics, N. Y., 1960; Fuller J. .Thompson W. R., Behavior genetics, N.Y. - L., 1960. Л. Г. Романова.
ГЕНЕТИКА РАСТЕНИЙ, раздел генетики, изучающий наследственность и изменчивость высших растений (генетич. исследования грибов и водорослей обычно относят к генетике микроорганизмов). Для генетич. изучения растений, кроме методов, к-рыми пользуются в др. областях генетики (в частности гибридологического анализа), применяют след, методы. С помощью моносо много анализа определяют роль каждой хромосомы в наследовании и развитии различных признаков растении. Этим методом (разработанным на дурмане) пользуются при изучении ряда аллополиплоидов (нек-рых пшениц, хлопчатника), а также диплоидов (ячменя). Большое значение в Г. р. приобретает экспериментальный мутагенез, к-рый даёт огромное разнообразие новых форм, используемых в селекции, и ценный материал для изучения генетики отд. видов растений. С помощью мутантов составляют генетические карты хромосом; на них исследуют действие изменённого гена (в гомо- и гетерозиготном состоянии) на развитие отд. признаков в разных условиях среды, на физиол. и биохим. особенности растений. Изучение мутантов способствует выяснению эволюции того или иного вида. К методам исследования эволюции растений относятся также гибридизация и анализ конъюгации хромосоме мей-озе у гибридов (неродств. хромосомы не конъюгируют). Важный метод - искусственный ресннтез существующих видов путём гибридизации и последующего удвоения числа хромосом (см. Полиплоидия). Значит, роль в эволюции растений, в т. ч. мн. культурных (пшеницы, овса, хлопчатника, картофеля, плодовых и др.), играет аллополиплоидия. После открытия действия алкалоида колхицина, препятствующего расхождению удвоившихся хромосом к разным полюсам клетки, для получения новых, иногда очень ценных форм широко используется автополиплоидия. Сочетая методы отдалённой гибридизации и цитогенетики, изучают роль отд. хромосом (и их участков) в наследовании признаков и разрабатывают приёмы, позволяющие получать вставки участков хромосом диких растений, обусловливающие развитие ценных признаков (напр., устойчивости к ржавчине), в хромосомы культурных растений. Роль ядра и цитоплазмы в наследовании и развитии признаков исследуют, применяя отдалённую гибридизацию и анализируя природу мужской цитоплазматич. стерильности, используемой при получении гетерозисных форм. В Г. р. широко исследуются апомиксис и явление самонесовместимости, т. е. неспособности растений к самооплодотворению, а также генетич. особенности растений само- и перекрёстноопылителей, вегетативно и апомиктически размножающихся форм. В Г. р. всё больше проникают идеи и методы молекулярной биологии (гибридизация ДНК, ДНК - РНК, изучение изозимов и др.). Методы популяционной генетики и биометрии применяют в Г. р. для разграничения генотипич. и паратипич. элементов в общей фенотипич. изменчивости признаков, что усиливает эффективность искусств, отбора. Все эти методы используют для улучшения хозяйственно ценных свойств с.-х. растений: урожайности, устойчивости к неблагоприятным условиям среды, ряда биохим. и технологич. особенностей растения (или его зерна), особенностей развития (озимость, яровость, раннеспелость и т. д.). Из высших растений генетически наиболее изучены кукуруза, арабидопсис (растение сем. крестоцветных, растительная дрозофила - модельный объект генетич. исследований), горох, томаты, ячмень. У этих растений методами гибридизации установлена локализация генов и составлены карты хромосом. Интенсивно изучается цитогепетика мягкой пшеницы - сложного 42-хромосомного аллополиплоида, возникшего в процессе эволюции при естеств. гибридизации трёх разных злаков с последующим удвоением числа хромосом у гибридов. Вклад Г. р. в селекцию огромен. Это, напр., использование гетерозиса в селекции кукурузы на основе мужской стерильности; введение высокоурожайным гибридам и сортам кормового ячменя генов, обеспечивающих высокое содержание лизина в зерне; создание низкорослых неполегающих высокоурожайных сортов пшеницы с использованием генов карликовости (-зелёная революция в Индии и др. странах); выведение урожайных и сахаристых триплоидных гибридов сахарной свёклы. Лит.: Вавилов Н. И., Избр. произв., т. 1, Л., 1967; Мичурин И. В., Соч., т. 1 - 4, М., 1948; Брюбейкер Дж. Л.,Сельскохозяйственная генетика, пер. с англ., М., 1966; Эллиот Ф., Селекция растений и цитогенетика, пер. с англ. ,М.. 1961: М юнтцинг А., Генетика, пер. с англ., М., 1967; Уильяме У., Генетические основы и селекция растений, пер. с англ., М., 1968; Цитогенетика пшеницы и ее гибридов, М., 1971; Генетические основы селекции растений, М., 1971. В.
В. Хвостова.
ГЕНЕТИКА ЧЕЛОВЕКА, отрасль генетики, тесно связанная с антропологией и медициной. Г. ч. условно подразделяют на антропогенетику, изучающую наследственность и изменчивость нормальных признаков человеческого организма, и генетику медицинскую, к-рая изучает его наследственную патологию (болезни, дефекты, уродства и др.). Г. ч. связана также с эволюционной теорией, т. к. исследует конкретные механизмы эволюции человека и его место в природе, с психологией, философией, социологией. Из направлений Г. ч. наи-бЪлее интенсивно развиваются цитогенетика, биохимич. генетика, иммуногенетика, генетика высшей нервной деятельности, физиологич. генетика. В Г. ч. вместо классич. гибридологического анализа применяют генеалогический метод, к-рый состоит в анализе распределения в семьях (точнее, в родословных) лиц, обладающих данным признаком (или аномалией) и не обладающих им, что раскрывает тип наследования, частоту и интенсивность проявления признака и т. д. При анализе семейных данных получают также цифры эмпирического риска, т. е. вероятность обладания признаком в зависимости от степени родства с его носителем. Генеалогич. методом уже показано, что более 1800 морфологич., биохимич. и др. признаков человека наследуется по законам Менделя (см. Генетика). Напр., тёмная окраска кожи и волос доминирует над светлой; пониженная активность или отсутствие нек-рых ферментов определяется рецессивными генами, а рост, вес, уровень интеллекта и ряд др. признаков-полимерными генами, т. е. системами из мн. генов. Мн. признаки и болезни человека, наследующиеся сцепленно с полом, обусловлены генами, локализованными в Х- или У-хромосоме. Таких генов известно ок. 120. К ним относятся гены гемофилии А н В, недостаточности фермента глюкозо-6-фосфат-дегидрогеназы, цветовой слепоты и др. Др. метод Г. ч.- близнецовый метод (см. Близнецы). Однояйцевые близнецы (ОБ) развиваются из одной яйцеклетки, оплодотворённой одним спермием; поэтому набор генов (генотип) у ОБ идентичен. Разнояйцевые близнецы (РБ) развиваются из двух и более яйцеклеток, оплодотворённых разными спермиями; поэтому их генотипы различаются так же, как у братьев и сестёр (сибсов). Сравнение внутрипарных различий между ОБ и РБ позволяет судить об относит, значении наследственности и среды в определении свойств человеческого организма. В близнецовых исследованиях особенно важен показатель конкордантности, выражающий (в % ) вероятность обладания данным признаком одним из членов пары ОБ или РБ, если его имеет другой член пары. Если признак детерминирован преим. наследств, факторами, то процент конкордантности намного выше у ОБ, чем у РБ. Напр., конкордантность по группам крови, к-рые детерминированы только генетически, у ОБ равна 100%. При шизофрении конкордантность у ОБ достигает 67%, в то время как у РБ - 12,1%; при врождённом слабоумии (олигофрении) - 94,5% и 42,6% соответственно. Подобные сравнения проведены в отношении ряда заболеваний. Т. о., исследования близнецов показывают, что вклад наследственности и среды в развитие самых разнообразных признаков различен и признаки развиваются в результате взаимодействия генотипа и внешней среды. Одни признаки обусловлены преим. генотипом, при формировании др. признаков генотип выступает в качестве предрасполагающего фактора (или фактора, лимитирующего норму реакции организма на действия внешней среды). Геном человека включает неск. миллионов генов, способных к тому же по-разному влиять на развитие признаков. В результате мутаций и перекомбинации генов возникает присущее человеку разнообразие по самым разным признакам. Гены человека мутируют каждый с частотой от 1 на 100 000 до 1 на 10 000 000 гамет на поколение. Распространение мутаций среди больших групп населения изучает популяционная Г. ч., позволяющая составить карты распространения генов, определяющих развитие нормальных признаков и наследственных болезней. Особый интерес для популяционной Г. ч. представляют изоляты - группы населения, в к-рых по к.-л. причинам (географич., экономич., социальным, религиозным и др.) браки заключаются чаще между членами группы. Это приводит к повышению частоты кровного родства вступающих в брак, а значит, и вероятности того, что рецессивные гены перейдут в гомозиготное состояние и проявятся, что особенно заметно при малочисленности изолята. Исследования в области Г. ч. продемонстрировали наличие естественного отбора в человеческих популяциях. Однако отбор у человека приобретает специфич. черты; он интенсивно действует только на эмбриональной стадии (т. н. самопроизвольные аборты - отражение такого отбора). Отбор в человеческом обществе осуществляется посредством дифференциальной брачности и плодовитости, т. е. в результате взаимодействия социальных и биологич. факторов. Мутационный процесс и отбор обусловливают огромное разнообразие (полиморфизм) по ряду признаков, присущее человеку, что делает его с биологич. точки зрения необычайно пластичным и приспособленным видом. Широкое использование в Г. ч. цитологич. методов способствовало развитию цитогенетики, где осн. объект исследования - хромосомы, т. е. структуры клеточного ядра, в к-рых локализованы гены. Установлено (1946), что хромосомный набор в клетках тела человека (соматических) состоит из 46 хромосом, причём женский пол определяется наличием двух Х-хромосом, а мужской - Х-хромосомы и Y-хромосомы. В зрелых половых клетках находится половинное (гаплоидное) число хромосом. Митоз, мейоз и оплодотворение поддерживают преемственность и постоянство хромосомного набора как в ряду клеточных поколений, так и в поколениях организмов. В результате нарушений указанных процессов могут возникать аномалии хромосомного набора с изменением числа и структуры хромосом, что приводит к возникновению т.н. хромосомных болезней, к-рые нередко выражаются в слабоумии, развитии тяжёлых врождённых уродств, аномалий половой дифференцировки или обусловливают самопроизвольные аборты. Успехи в развитии Г. ч. сделали возможными предупреждение и лечение наследственных заболеваний. Один из эффективных методов их предупреждения - медико-генетическое консультирование с предсказанием риска появления больного в потомстве лиц, страдающих данным заболеванием или имеющих больного родственника. Достижения биохимич. Г. ч. раскрыли первопричину (молекулярный механизм) мн. наследственно обусловленных дефектов, аномалий обмена веществ, что способствовало разработке методов экспресс-диагностики, позволяющих быстро и рано выявлять больных, и лечения мн. прежде неизлечимых наследств, болезней. Чаще всего лечение состоит во введении в организм веществ, не образующихся в нём вследствие генетич. дефекта, или в составлении специальных диет, из к-рых устранены вещества, оказывающие токсич. действие на организм в результате наследственно обусловленной неспособности к их расщеплению. Мн. генетич. дефекты исправляются с помощью своеврем. хирургич. вмешательства или педагогич. коррекции. Практич. мероприятия, направленные на поддержание наследств, здоровья человека, на охрану генофонда человечества, осуществляются через систему медико-генетических консультаций. Осн. цель медико-генетич. консультирования - информировать заинтересованных лиц о вероятности риска появления в потомстве больных. К медико-генетич. мероприятиям относится также пропаганда генетич. знаний среди населения, т. к. это способствует более ответственному подходу к деторождению. Медико-генетич. консультация воздерживается от мер принудительного или поощрительного характера в вопросах деторождения или вступления в брак, принимая на себя лишь функцию информации. Большое значение имеет система мер, направленных на создание наилучших условий для проявления положит, наследств, задатков и предотвращение вредных воздействий среды на наследственность человека. Г. ч. представляет собой естественнонаучную основу борьбы с расизмом, убедительно показывая, что расы - это формы адаптации человека к конкретным условиям среды (климатич. и иным), что они отличаются друг от друга не наличием хороших или плохих генов, а частотой распространения обычных генов, свойственных всем расам. Г. ч. показывает, что все расы равноценны (но не одинаковы) с биологич. точки зрения и обладают равными возможностями для развития, определяемого не генетич., а социально-историч. условиями. Констатация биологич. наследственных различий между отд. людьми или расами не может служить основанием для к.-л. выводов морального, юридического или социального порядка, ущемляющих права этих людей или рас (см. Геноцид, Сегрегация). Лит.: Ниль Дж. и Шэлл У., Наследственность человека, пер. с англ., М., 1958; К а н а е в И. И., Близнецы, М.- Л., 1959; Штерн К., Основы генетики человека, пер. с англ., М., 1965; Маккьюсик В., Генетика человека, пер. с англ., М., 1967; Биология человека, пер. с англ., М., 1968; Эфроимсон В. П., Введение в медицинскую генетику, 2 изд., М., 1968; Основы цитогенетики человека, [М., 1969]; Li Ching-chun, Human genetics, N. Y., 1961. К.
Н. Гринберг, А. А. Прокофьева-Белъговская.
ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ, вероятностные процессы, определяющие изменение частоты разных аллелей в популяции. В больших, свободно скрещивающихся популяциях в отсутствие отбора и давления мутаций соотношение аллелей, независимо от их абсолютной исходной частоты, должно сохраняться во всех поколениях. Однако в реальных, ограниченных по численности популяциях частота генов не остаётся постоянной не только под давлением мутаций и отбора, но и в силу случайных отклонений. Детальный анализ Г.-а. п. был проведён сов. генетиками Н. П. Дубининым (1931), Н. П. Дубининым и Д. Д. Ромашовым (1932), англ.- Р. Фишером (1931) и амер.- С. Райтом (1931). Случайные колебания частоты аллелей в популяции связаны с тем, что распределение аллелей между гаметами и комбинирование гамет в зиготе - вероятностные процессы. Г.-а. п. оказывают несистематич. эффект, т. к. частота аллелей в разных поколениях может повышаться или понижаться. В малых популяциях или в популяциях, к-рые распадаются под действием изоляционных механизмов на отдельные подгруппы, может происходить чисто случайная стабилизация аллелей (гомозиготы) или их элиминация; в результате довольно быстро проявляются новые стабилизированные комбинации генов. Наиболее отчётливо Г.-а. п. проявляются при возникновении новых изолированных популяций. Напр., в секте мен-нонитов (Ланкастер, штат Пенсильвания, США), насчитывающей ок. 8000 человек, значителен процент карликов с многопа-лостью (13% меннонитов гетерозиготны по гену, к-рый в гомозиготном состоянии обусловливает появление таких карликов); это объясняется тем, что члены секты вступают в брак только между собой, а такая изоляция способствует появлению гомозиготных индивидуумов. В больших популяциях Г.-а. п. не могут обусловить такой стабилизации или элиминации аллелей, т. к. влияние этих процессов компенсируется за счёт разных факторов в последующих поколениях или в разных подразделениях популяции. Теория Г.-а. п. объяснила генетич. последствия изоляции, судьбу рецессивных мутаций на уровнях малых концентраций и эволюцию популяций по нейтральным признакам. Г.-а. п. объясняют мн. расовые различия человека, возникшие без действия отбора. Наряду с термином Г.-а. п. широко используется термин дрейф генов, предложенный С. Райтом. Сов. генетик С. С. Четвериков, подчёркивая роль вероятностно-статистич. закономерностей при дрейфе генов, предлагал назвать это явление генетико-стохастич. процессами. Лит: Дубинин Н. П., Эволюция популяции и радиация, М., 1966, с. 421 - 33. Н.
П. Дубинин, В. Н. Сойфер.
ГЕНЕТИКОВ И СЕЛЕКЦИОНЕРОВ ОБЩЕСТВО Всесоюзное имени Н. И. Вавилова (ВОГИС), научно-общественная организация при АН СССР, объединяющая учёных и практиков СССР, работающих в области генетики и селекции. Создано в 1965. 30-31 мая 1966 в Москве состоялся учредительный съезд общества; был утверждён устав общества, избраны центральный совет (80 чел.) и президиум (23 чел.; находится в Москве). Президентом избран Б. Л. Астауров. К началу 1971 общество насчитывало 3670 чл., имело 26 отделений в столицах союзных республик, краевых, областных центрах и др. городах. Цели и задачи ВОГИС: активное участие в развитии всех отраслей генетики и селекции, повышение квалификации членов общества и реализация их исследований, популяризация и пропаганда новейших теоре-тич. и практич. достижений в области генетики и селекции, содействие преподаванию генетики и селекции в средней и высшей школе. В.
Ф. Мирек.
ГЕНЕТИЧЕСКАЯ ИНФОРМАЦИЯ, заложенная в наследственных структурах организмов (в хромосомах, цитоплазме, клеточных органеллах), получаемая от предков в виде совокупности генов информация о составе, строении и характере обмена составляющих организм веществ (прежде всего белков и нуклеиновых кислот) и связанных с ними функциях. У многоклеточных форм при половом размножении Г. и. передаётся из поколения в поколение через посредство половых клеток - гамет, единственная функция к-рых - передача и хранение Г. и. У микроорганизмов и вирусов имеются особые типы передачи Г. и. (см. Сексдукция, Трансдукция, Трансформация). Г. и. заключена преим. в хромосомах, где она зашифрована в определённой линейной последовательности нуклео-тидов в молекулах дезоксирибонуклеи-новой кислоты - ДНК (см. Генетический код). Г. и. реализуется в ходе онтогенеза - развития особи -передачей Г. и. от гена к признаку. Все клетки организма возникают в результате делений единственной исходной клетки - зиготы - и потому имеют один и тот же набор генов - потенциально одну и ту же Г. и. Специфичность клеток разных тканей определяется тем, что в них активны разные гены, т. е. реализуется не вся Г.и., а только её часть, необходимая для функционирования данной ткани. Ю. С.
Дёмин.
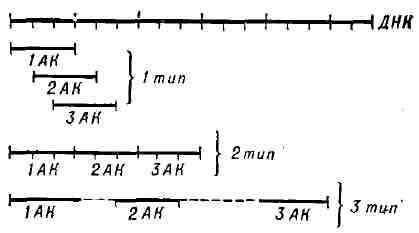
ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ, схемы относительного расположения сцепленных между собой наследств, факторов - генов. Г. к. х. отображают реально существующий линейный порядок размещения генов в хромосомах (см. Цитологические карты хромосом) и важны как в теоретич. исследованиях, так и при проведении селекционной работы, т. к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Г. к. х., можно по наследованию сигнального гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; напр., ген, определяющий сморщенный эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения. Многочисленные факты отсутствия (вопреки Менделя законам) независимого распределения признаков у гибридов второго поколения были объяснены хромосомной теорией наследственности. Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество к-рых, т. о., соответствует у каждого организма гаплоидному числу хромосом (см. Гаплоид). Амер. генетик Т. X. Морган показал, однако, что сцепление генов, расположенных в одной хромосоме, у диплоидных организмов (см. Диплоид) не абсолютное; в нек-рых случаях перед образованием половых клеток между однотипными, или гомологичными, хромосомами происходит обмен соответств. участками; этот процесс носит назв. перекреста, или кроссинговера. Обмен участками хромосом (с находящимися в них генами) происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации). Генетич. анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Г. к. х. выражают как % кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему кол-ву изученных особей); единица этого расстояния - морганида - соответствует частоте кроссинговера в 1 % . Г. к. х. составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, номере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые назв. мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Г. к. х. можно только для объектов, у к-рых изучено большое число мутантных генов. Напр., у дрозофилы идентифицировано св. 500 генов, локализованных в её 4 группах сцепления, у кукурузы - ок. 400 генов, распределённых в 10 группах сцепления (рис. 1). У менее изученных объектов числэ обнаруженных групп сцепления меньше гаплоидного числа хромосом. Так, v домовой мыши выявлено ок. 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом. У бактерий, к-рые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления (рис. 2). При переносе генетич. материала из клетки-донора в клетку-реципиент, напр, при конъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110- 120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбииантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Г. к. х. бактерий, детально разработанных у ряда видов. Ещё более детализированы Г. к. х. нек-рых бактериофагов. См. также Генетика, Мутация. Рис. 1. Генетические карты 7 -10-й хромосом кукурузы. Цифры по длине хромосом обозначают расстояние от конца хромосомы в морганидах; буквы - сокращённые названия признаков , определяемых соответствующими генами. Рис. 2. Генетическая карта хромосомы тсишечной палочки (Eschenchia coli К 12). Цифры означают время (в мин), необходимое для переноса в клетку-реципиент генетических маркёров, контролирующих биосинтез ряда аминокислот, а также устойчивость к стрептомицину и к фагу Т6 эти цифры характеризуют расстояние между генами. Обозначения: ade - аденин; his - гистидин; try - триптофан; gal - галактоза; lac - лактоза; pro - пролин; leu - лейцин; tre - треонин; met - метионин; агд - аргинин; mt - маннит; xyl - ксилоза; mal - мальтоза; ser - серии; gly - глицин; sir иТ6, - устойчивость к стрептомицину или фагу Т6. Лит,.: Лобашев М. Е., Генетика, 2 изд., Л., 1967; Медведев Н. Н., Практическая генетика, 2 изд., М., 1968; Актуальные вопросы современной генетики. Сб. ст., М., 1966; Жакоб Ф., Вольман Э., Пол и генетика бактерий, пер. с англ., М., 1962; Бензер С., Тонкая структура гена, в сб.: Молекулярная генетика, пер. с англ., М., 1963; Xэис У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Рейвин А. У., Эволюция генетики, пер. с англ., М., 1967; Мюнтцинг А., Генетика, пер. с англ.,2 изд., М., 1967; Уотсон Д ж., Молекулярная биология гена, пер. с англ., М., 1967. В. С.
Андреев.
ГЕНЕТИЧЕСКИЕ
РЯДЫ, группы органич. соединений
с одинаковым числом атомов углерода в молекуле, различающихся функциональными
группами; см. Гомологические ряды.
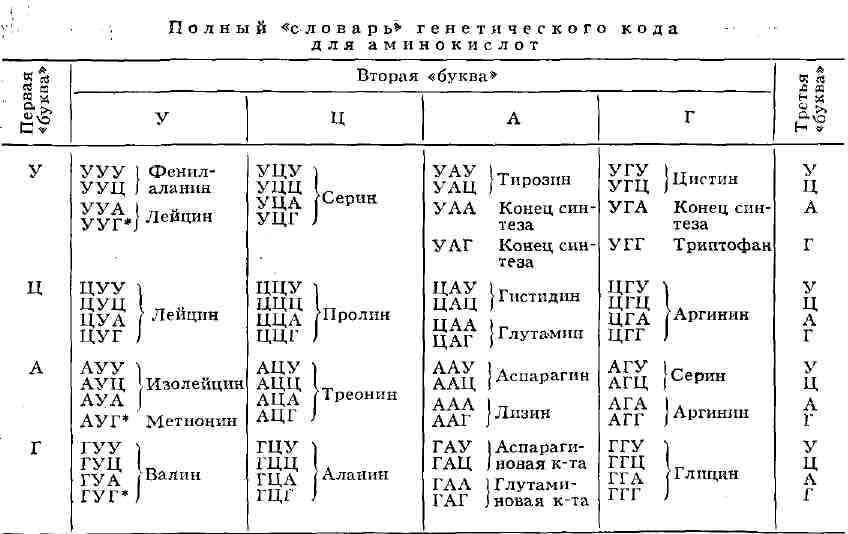
ГЕНЕТИЧЕСКИЙ АНАЛИЗ, совокупность методов изучения наследственных свойств организмов. Г. а. включает: 1) Гибридологический метод, изучающий законы наследственности, а также строение и поведение наследств, структур с помощью спец. видов скрещиваний (см. Гибридологический анализ). 2) Цитогенетический метод, развившийся на стыке генетики и цитологии. Главная его задача - установление связи между закономерностями наследования и строением и функциями хромосом (составление цитологических карт хромосом, геномный анализ и др.). 3) Молекулярно-генетический метод, получивший развитие в связи с новыми биохим. и физ.-хим. методами анализа наследственных структур. С его помощью изучается связь между молекулярным строением генов и синтезируемыми в соответствии с заложенной в них информацией белками. Лит.: Серебровский А. С., Генетический анализ, М., 1970 (библ.). Ю.С.Дёмин. ГЕНЕТИЧЕСКИЙ КОД, система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов. В природных нуклеиновых кислотах - дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК)-встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой к-те), различающихся по входящему в их состав азотистому основанию (см. Пуриновые основания, Пиримидиновые основания). В ДНК встречаются основания: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых к-т обнаружено ок. 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных Сахаров. Т. к. кол-во кодирующих знаков Г. к. (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. кол-во нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. Различных сочетаний по 2 нуклеотида возможно лишь 42 = 16, но этого также недостаточно для зашифровки всех аминокислот. Амер. учёный Г. Гамов предложил (1954) модель триплетного Г. к., т. е. такого, в к-ром 1 аминокислоту кодирует группа из трёх нуклеотидов, наз. кодоном. Число возможных триплетов равно 43 = 64, а это более чем втрое превышает число распространённых аминокислот, в связи с чем было высказано предположение, что каждой аминокислоте соответствует неск. кодонов (т. н. вырожденность кода). Было предложено много различных моделей Г. к., из к-рых серьёзного внимания заслуживали три модели (см. рис.): перекрывающийся код без запятых, неперекрывающийся код без запятых и код с запятыми. В 1961 Ф. Крик (Великобритания) с сотрудниками получил подтверждение гипотезы триплетного неперекрывающегося кода без запятых. Установлены след. осн. закономерности, касающиеся Г. к.:
Модели генетического кода:
1-й тип - перекрывающийся код без запятых; 2-й тип - неперекрывающийся
код без запятых; 3-й тип - код с "промежутками", т. е. код с запятыми.
1) между последовательностью нуклеотидов и кодируемой последовательностью аминокислот существует линейное соответствие (колинеарность Г. к.); 2) считывание Г. к. начинается с определённой точки; 3) считывание идёт в одном направлении в пределах одного гена; 4) код является неперекрывающимся; 5) при считывании не бывает промежутков (код без запятых); 6) Г. к., как правило, является вырожденным, т. е. 1 аминокислоту кодируют 2 и более триплетов-синонимов (вырожденность Г. к. уменьшает вероятность того, что мутационная замена основания в триплете приведёт к ошибке); 7) кодовое число равно трём; 8) код в живой природе универсален (за нек-рыми исключениями). Универсальность Г. к. подтверждается экспериментами по синтезу белка in vitro. Если в бесклеточную систему, полученную из одного организма (напр., кишечной палочки), добавить нуклеиновокислотную матрицу, полученную из др. организма, далеко отстоящего от первого в эволюционном отношении (напр., проростков гороха), то в такой системе будет идти белковый синтез. Благодаря работам амер. генетиков М. Ниренберга, С. Очоа, X. Корана известен не только состав, но и порядок нуклеотидов во всех ко донах (см. табл., построенную по данным опытов с кишечной палочкой). Из 64 кодонов у бактерий и фагов 3 ко-дона - УАА, УАГ и УГА - не кодируют аминокислот; они служат сигналом к освобождению полипептидной цепи с рибосомы, т. е. сигнализируют о завершении синтеза полипептида. Их наз. терминирующими кодонами. Существуют также 3 сигнала о начале синтеза - это т. н. инициирующие кодоны - АУГ, ГУГ и УУГ,- к-рые, будучи включёнными в начале соответствующей информационной РНК (и-РНК), определяют включение формилметионина в первое положение синтезируемой полипептидной цепи. Приведённые данные справедливы для бактериальных систем; для высших организмов многое ещё не ясно. Так, кодон УГА у высших организмов может быть значащим; не совсем понятен также механизм инициации полипептида. Реализация Г. к. в клетке
происходит в два этапа. Первый из них протекает в ядре; он носит назв.
транскрипции и заключается в синтезе молекул и-РНК на соответствующих участках
ДНК. При этом последовательность нуклеотидов ДНК "переписывается" в нуклеотидную
последовательность РНК. Второй этап - трансляция - протекает в цитоплазме,
на рибосомах; при этом последовательность нуклеотидов и-РНК переводится
в последовательность аминокислот в белке; этот этап протекает при участии
транспортной РНК (т-РНК) и соответствующих ферментов (см. Белки, раздел
Биосинтез).
В начале цепи и-РНК данный кодон определяет начало синтеза полипептидной цепи и кодирует аминокислоту формилметионин. От готовых полиплоидных цепей формильная группа или вся аминокислота может быть отщеплена с помощью соответствующих ферментов. Лит.: Общая природа генетического
кода для белков, в сб.: Молекулярная генетика, пер. с англ., М. 1963; Крик
Ф., Генетический код (I), в кн.: Структура и функция клетки, пер. с англ.,
М., 1964, с. 9 - 23; Н и-ренберг М., Генетический код (II), там же, с.
24 - 41; Xэй с У., Генетика бактерий и бактериофагов, пер. с англ., М.,
1965; Хартман Ф.,Саскайнд З., Действие гена, пер.сангл., М., 1966; Бреслер
С.Е., Введение в молекулярную биологию, 2 изд., М. - Л., 1966; Ингрэм В.,
Биосинтез макромолекул, пер. с англ., М., 1966; Лобашев М. Е., Генетика,
2 изд., Л., 1967; Уотсон Дж., Молекулярная биология гена, пер. с англ.,
М., 1967; Сойфер В. Н., Молекулярные механизмы мутагенеза, М., 1969; Дубинин
Н. П., Общая генетика, М., 1970. Н.П.Дубинин, В.Н.Сойфер.
ГЕНЕТИЧЕСКИЙ ПОЛИМОРФИЗМ,сосуществование в пределах популяции двух или нескольких различных наследственных форм, находящихся в динамич. равновесии в течение неск. и даже мн. поколений. Чаще всего Г. п. обусловливается либо варьирующими давлениями и векторами (направленностью) отбора в различных условиях (напр., в разные сезоны), либо повышенной относит, жизнеспособностью гетерозшот. Один из видов Г. п.- сбалансированный Г. п. - характеризуется постоянным оптимальным соотношением полиморфных форм, отклонение от к-рого оказывается неблагоприятным для вида, и автоматически регулируется (устанавливается оптимальное соотношение форм). В состоянии сбалансированного Г. п. у человека и животных находится большинство генов. Различают неск. форм Г. п., анализ к-рых позволяет определять действие отбора в природных популяциях. Лит.:
Тимофеев-Ресовский Н. В., Свирежев Ю. М., О генетическом полиморфизме
в популяциях, Генетика, 1967, № 10.
ГЕНЕТИЧЕСКОЕ ДЕЙСТВИЕ ИЗЛУЧЕНИЙ, радиационный мутагенез, возникновение наследственных изменений (мутаций) при облучении организмов. Г. д. и.- важная часть биологического действия ионизирующих излучений, исследуемая радиационной генетикой. Первые стабильные радиорасы у дрожжей получены сов. биологами Г. А. Надсоном и Г. С. Филипповым (1925); данные о повышении частоты мутаций у дрозофилы при рентгеновском облучении опубликованы амер. генетиком Г. Меллером (1927). Мутагенный эффект вызывают все типы ионизирующих излучений, а также ультрафиолетовые лучи, если их действию подвергаются наследственные структуры любых организмов - от вирусов и бактерий до высокоорганизованных животных, включая человека; при этом у сложных организмов мутации могут возникать как в половых клетках - гаметах, так и в клетках тела - соматических. Облучение может вызывать все типы мутаций (генные, хромосомные, геномные и цито-плазматические). В определ. интервале доз частота мутаций возрастает пропорционально дозе облучения; при увеличении дозы выше нек-рого значения линейность кривых, описывающих зависимость частоты мутаций от дозы, нарушается. Вновь возникающие мутации являются обычно рецессивными (см. Рецессивность) и вредными. Повышение радиоактивного фона ведёт к накоплению в популяциях организмов, в т. ч. и человека, скрытых вредных мутаций. Важное практич. применение Г. д. и.- радиационная селекция, т. е. отбор хоз.-ценных мутаций, получаемых гл. обр. у культурных растений и пром. микроорганизмов в результате их облучения. Выведенные таким способом новые сорта овса, ячменя, гороха, арахиса, плодовых и декоративных культур и др. уже занимают большие посевные площади. Мн. высокопродуктивные пром. штаммы микроорганизмов - продуцентов антибиотиков, витаминов, аминокислот-также получены путём радиационного мутагенеза. Лит.: Итоги науки. Биологические науки, т. 3-Ионизирующие излучения и наследственность, М., 1960; Астауров Б. Л., Функциональный принцип в оценке относительной значимости радиационных поражений ядра н цитоплазмы, в сб.: Первичные механизмы биологического действия ионизирующих излучений, М., 1963; Ли Д. Э., Действие радиации на живые клетки, пер. с англ., М., 1963; Алнханян С. И., Селекция промышленных микроорганизмов, М., 1968; Тимофеев-Ресовский Н. В., Иванов В. И., Корогодин В. И., Применение принципа попаданий в радиобиологии, М., 1968. Н.
В. Тимофеев-Ресовский, В. И. Иванов.
ГЕНЕТТА (Genetta), род хищных млекопитающих сем. виверровых. Длинное (до 55 см), приземистое и необычайно гибкое тело покрыто короткой, довольно грубой шерстью; окраска пятнистая; хвост не пушистый, до 50 см', у его основания расположены железы, выделяющие резко пахнущую жидкость-мускус. 6 видов; распространены гл. обр. в саваннах и тропич. лесах Африки. Обыкновенная Г. (G. genetta) широко распространена по всей Африке, встречается и в Юго-Зап. Европе (Испания, Франция), где населяет лесистые и безлесные горы и низменности, обитая преим. около водоёмов. По повадкам Г. напоминает хорьков. Питается мелкими зверьками, птицами и их яйцами, а также беспозвоночными. Иногда вредит птицеводству. Ведёт преим. ночной образ жизни. Г. легко приручаются; в Африке их иногда содержат дома для истребления крыс и мышей. Лит.: Mammals
of the world, v. 2, Bait., 1964.
ГЕНЗЕЛЬТ Адольф Львович (12.5.1814, Швабах, Бавария,-10.10.1889, Вармбрунн, Силезия), русский пианист, педагог и композитор. Род. в нем. семье. В 1836 начал концертную деятельность в Берлине. С 1838 жил в Петербурге, занимаясь преим. преподаванием игры на фортепьяно (среди его учеников - В. В. Стасов, И. Ф. Нейлисов, Н. С. Зверев). В 1872-75 редактор муз. журнала Нувеллист; в 1887-88 профессор Петерб. консерватории. Игра Г. отличалась тонкой художеств. выразительностью, тщательной отделкой деталей и замечательным технич. мастерством. Фортепьянные произв. Г. (св. 40 опусов) мелодичны, изящны (особенно выделяются этюды, блестяще разработанные в пианистич. отношении), но подчас носят налёт салонной виртуозности. Г. принадлежит множество редакций, переложений и обработок для фортепьяно (в т. ч. обработки рус. нар. песен и произв. рус. композиторов). Автор инструктивно-педагогич. сочинения На многолетнем опыте основанные правила преподавания фортепианной игры (1868). Лит.: Ф.
[Финдеизен Н.], Адольф Гензельт, Русская музыкальная газета, 1899, № 37;
Алексеев А., Русские пианисты. Очерки и материалы по истории пианизма,
в. 2, М.- Л., 1948; Музалевский В., Русская фортепианная музыка. Очерки
и материалы по истории русской фортепианной культуры, Л.- М., 1949.
ГЕНИАЛЬНОСТЬ, наивысшая степень проявления творческих сил человека. Термин Г. употребляется как для обозначения способности человека к творчеству, так и для оценки результатов его деятельности. Предполагая врождённую способность к продуктивной деятельности в той или иной области, гений, в отличие от таланта, представляет собой не просто высшую степень одарённости, а связан с созданием качественно новых творений, открытием ранее неизведанных путей творчества. Деятельность гения реализуется в определённом историч. контексте жизни человеческого общества, из к-рой гений черпает материал для своего творчества. В психологии творчества Г. изучается под углом зрения индивидуальных особенностей личности (её психич. склада, способностей и т. д.), а также разнообразных факторов, влияющих на творчество. С психологич. точки зрения гений не может рассматриваться как особый тип личности. Разнообразные попытки выделить к.-л. общие психологич. или психопатологич. черты Г. оказались неудовлетворительными: гениальные люди обнаруживают значительные индивидуальные различия с точки зрения одарённости, характера, культуры, интересов, навыков и т. д. Сам творческий процесс гения также принципиально не отличается с психологич. точки зрения от творческого процесса других одарённых людей. В ряде концепций Г., особенно начиная с Ч. Ломброзо, постулировалась связь между Г. и психич. расстройством, что, однако, не находит всеобщего подтверждения: дисгармония душевной жизни или предрасположенность к ней, свойственная многим гениальным людям, не является неизменным спутником Г. Историч. воззрения на природу Г. и её оценка связаны с общим пониманием творческого процесса (см. Творчество). Так, от античности идёт взгляд на Г. как род иррационального вдохновения, божественного наития (Платон, неоплатонизм). Начиная с эпохи Возрождения (Леонардо да Винчи, Дж. Вазари, Скалигер) получает распространение культ гения как творч. индивидуальности, достигающий своего апогея в период романтизма (предромантич. течение Бури и натиска в Германии, романтизм и вышедшие из него учения Т. Кар-лейля, Ф. Ницше с характерным для них противопоставлением гения и массы). В 18 в. складывается понятие гения в совр. смысле этого слова, к-рое у А. Шефтсбери становится одним из основных эстетич. понятий (гений творит подобно силе природы; его создания оригинальны в отличие от подражающего художника). И. Кант также подчёркивает оригинальность и естественность творчества гения: гений - это ...прирожденные задатки души..., через которые природа дает искусству правило (Соч., т. 5, М., 1966, с. 323). Ф. Шиллер раскрывает природу Г. через понятие наивности как инстинктивного следования безыскусств. природе и как способности к непредвзятому постижению мира. В 19-20 вв. получают развитие психологич. (в т. ч. психиатрические), со-циально-психологич., а также социология, исследования различных аспектов Г. и творчества. Лит.: Грузенберг
С. О., Гений и творчество, Л., 1924; Жоли Г., Психология великих людей,
СПБ, 1894; Оствальд В., Великие люди, [пер. с нем.], СПБ, 1910; W о 1 f
H.. Versuch
einer Geschichte des Geniebegriffes, Hdlb.,1923; Genetic studies of genius,
ed. L. M. Terman, 2 ed., v.i -4, [Stanford], 1926-47; Kretschmer E., Geniale
Menschen, 5Aufl.,B., 1958.
ГЕНИЙ (лат. genius), 1) в др.-рим. религии и мифологии добрый дух, сверхъестественное существо, охраняющее человека на протяжении всей его жизни. День рождения римлянина считался праздником его Г., к-рому приносились жертвы, совершались возлияния и курения. Римляне почитали также Г.- покровителей семьи, общины, города и всего римского народа. Культ Г. в Др. Риме - разновидность широко распространённого у многих народов в период разложения общинно-родовых отношений культа личных духов-покровителей. 2) Высшая
степень творческой одарённости, а также человек, обладающий подобной одарённостью;
см. Гениальность.
ГЕНИСАРЕТСКОЕ
ОЗЕРО, одно из названий Тивериадского
озера, расположенного в Западной Азии.
ГЕНИЧЕСК, город,
центр Генического р-на Херсонской обл. УССР. Порт на Азовском м. Ж.-д.
ст. 20 тыс. жит. (1970). Рыбоконсервный, винодельческий, арматурный, стройматериалов
заводы. Мед. училище. Добыча соли.
ГЕНИЧЕСКИЙ
ПРОЛИВ, Тонкий пролив между
Арабатской стрелкой и берегом, соединяет Азовское м. с зал. Сиваш. Дл.
ок. 4 км, шир. 80-150 м. Глуб. 4,6 м. Течения зависят
от ветров. На берегу - порт Геническ.
ГЕНК
(Genk), город в Бельгии, в пров. Лимбург. 56 тыс. жит. (1969). Добыча угля
(копи Винтерслаг), металлургич., хим. промышленность. Автомобильный завод.
ГЕНКИН Дмитрий Михайлович (19.9. 1884, Калуга,-24.1.1966, Москва), советский юрист, засл. деят. науки РСФСР (1945), доктор юридич. наук (1940). Чл. КПСС с 1964. Специалист в области гражд. права. С 1909 на научной и преподават. работе, в 1919 проф. Ин-та нар. х-ва им. Г. В. Плеханова (в 1919-21 ректор этого ин-та); проф. Всесоюзной академии внешней торговли, пред. Внешнеторговой арбитражной комиссии. Автор научных трудов, гл. обр. по теории гражд. права, правовым вопросам внешней торговли, правовому регулированию деятельности пром. предприятий. Награждён орденом Ленина, 3 др. орденами, а также медалями. Соч.:
Законодательство о промысловой кооперации, М., 1930; Недействительность
сделок, совершенных с целью, противной закону, Уч. зап. Всесоюзного ин-та
юридических наук, 1947, в. 5; Право собственности в СССР, М., 1961; Правовые
вопросы хозяйственного расчета государственных промышленных предприятий,
М., 1966.
ГЕННАДИ Григорий Николаевич [18(30). 3.1826, Петербург,-26.2(9.3). 1880, там же], русский библиограф. Библиографич. деятельность начал в 1849. До кон. 50-х гг. занимался библиографией самых различных, не связанных между собой отраслей. С 60-х гг. работал в основном над Справочным словарем о русских писателях и учёных. Крупнейшие труды Г.: Литература русской библиографии (1858) - систе-матич. аннотированный указатель рус. библиографич. лит-ры, и Справочный словарь о русских писателях и учёных, умерших в XVIII и XIX столетиях, и список русских книг с 1725 по 1825 [т. 1-3 (буквы А - Р), Берлин - М., 1876-1908, т. 4 остался в рукописи], содержащий сведения о рус. писателях и учёных, умерших до 1874, а также о рус. анонимных книгах, изданных в 1725- 1825. Г. был одним из первых библиографов Пушкина, редактировал два изд. его соч., выпущенных Я. А. Исаковым (1859-60, 1869-71); в 1-м изд. Г. в качестве приложения опубликовал Библиографический список всех произведений А. С. Пушкина... (1860). Лит.: И
вас к У. Г., Г. И. Геннади. Обзор жизни и трудов, М., 1913.
ГЕННАДИЙ (г. рожд. неизв.-ум. 1505), русский церк. и политический деятель. Назначенный в 1484 новгородским архиепископом, Г. должен был стать проводником великокняж. политики в недавно присоединённом к Русскому централизованному гос-ву Новгороде. Но вскоре у Г. о вел. кн. Иваном III обнаружились противоречия из-за земельных владений архиепископской кафедры. Вместе с видным церковным деятелем Иосифом Волоцким Г. боролся против новгородско-моск. ереси (жидовствующих) и жестоко расправлялся с еретиками (см. Ереси). Публицистическая и переводческая деятельность лит. кружка, организованного Г., была связана с борьбой против еретиков. Обеспокоенный и смущённый малой грамотностью своих сподвижников и образованностью вольнодумцев, Г. выступал за создание училищ для духовенства и распространял церк.-полемич. и астрономич. лит-ру. При дворе Г. было написано Слово кратко в защиту церк. имуществ (первонач. название-Собрание на лихоимцев) и составлен первый в России полный библейский кодекс - ГеНнадиевская библия (1499). В 1504 Г. и его сподвижникам удалось добиться полного осуждения еретиков, но сам Г. по воле вел. князя был смещён с архиепископской кафедры. Умер в опале. Лит.: Слово кратко в защиту монастырских имуществ, Чтения в обществе истории и древностей российских, 1902, кн. 2, отд. 2; Собрание на лихоимцев - неизданный памятник русской публицистики конца XV в., Тр. Отдела древнерусской литературы, т. 21, М.- Л., 1965; Лурье Я. С.. Идеологическая борьба в русской публицистике конца XV - начала XVI вв., М.- Л., 1960. Я.
С. Лурье.
ГЕННЕГАУ (флам.
Henegouwen, франц. Hainaut), ср.-век. графство, затем провинция ср.-век.
Нидерландов, с 1830 - Бельгии; см. Эно.
ГЕННЕП (van Gennep) Арнольд ван (23.4.1873, Людвигсбург,- 7.5.1957, Эпер-не), французский этнограф, фольклорист, исследователь первобытной религии. Президент общества франц. этнографов (1952-57), основатель ряда этнографич. изданий. Автор многочисл. трудов по общей этнографии и этнографии Франции; первым из франц. этнографов применил метод этнографич. картографирования. Соч.:
Religions, moeurs et legendes. Essais d'ethnoaraphie et de linguistique,
v. 1-5, P., 1908-14; Le folklore, P., 1924; Manuel de folklore francais
contemporaine, pt. 1, 3, 4, P.. 1937-58.
ГЕННИН, де Геннин Виллим Иванович (Георг Вильгельм) [11(21).10. 1676-12(23).4.1750], специалист и организатор горного и металлургического производства в России, генерал-лейтенант. Голландец по происхождению; с 1698 на рус. службе. Участвовал в Северной войне 1700-21. В 1713-22 был нач. горных заводов Олонецкого края. С 1722 нач. уральских горных заводов. При нём усовершенствованы старые заводы и выстроено 9 новых. После возвращения с Урала (1734) был управляющим Гл. арт. канцелярии, перестраивал тульские з-ды, заведовал сестрорецкими з-дами. Соч.: Описание уральских и сибирских заводов, М., 1937. Лит.: Павленко
Н. И., Развитие металлургической промышленности России в первой половине
XVIII в., М., 1953.
ГЕНОВ (псевд. Цонев) Гаврил Димитров (1.2.1892, с. Живовци, Михайловградский округ,-20.1.1934, Москва), деятель болг. рабочего движения. Сын крестьянина-бедняка. Работал сел. учителем, принимал участие (с 1912) в деятельности Видинской учительской орг-ции Болг. с.-д. партии (тесных социалистов). Во время Балканских войн 1912-13 и 1-й мировой войны 1914-18-офицер; за ан-тивоен. деятельность подвергался репрессиям. В 1920-23 секретарь Врачанского окружкома компартии. Во время Сентябрьского антифашистского восстания 1923 возглавлял вооружённую борьбу во Врачанском округе, был чл. Гл. революц. к-та. После поражения восстания эмигрировал в Югославию. В 1925- 1926 чл. заграничного бюро ЦК БКП (тесных социалистов). С 1927 - в СССР, работал в Крестьянском интернационале и болг. секции ИККИ. Окончил междунар. Ленинскую школу. Редактировал перевод соч. В. И. Ленина на болг. язык. Соч.: Сентябрьское восстание в Болгарии 1923 года, М.- Л.,1934 (совм. с А. Владимировым). М.
А. Бирман.
ГЕНОГЕОГРАФИЯ, направление исследований в пограничной между генетикой и биогеографией области, сформулированное А. С. Серебровским в 1928- 1929. Осн. задача Г.- установление геогр. распространения и, по возможности, частот аллелей, определяющих осн. признаки и свойства в пределах всего или части ареала • изучаемого вида организмов. Г. изучает также причины распространения аллелей. Проведение гено-геогр. работ возможно лишь у тех видов, у к-рых в экспериментально-генетич. исследованиях установлена связь между изучаемыми признаками и генами. Шире возможности т. н. феногеографин, изучающей геогр. распространение элементарных признаков в пределах ареала вида. Г. (и феногеография) имеет большое теоретич. значение в исследованиях по систематике и эволюции видов. Практич. и прикладное значение Г. имеет в установлении генофондов домашних животных и культурных растений как одной из основ породного и сортового районирования и селекции, а также в генетике человека и, особенно, в мед. генетике. Лит.: Серебровский А. С., Геногеография и генофонд сельскохозяйственных живвтных СССР, Научное слово, 1928, № 9; е г о ж е. Проблемы и метод геногеографии, в кн.: Труды Всесоюзного съезда по генетике, селекции, семеноводству и племенному животноводству, т. 2, Л., 1930. Н.
В. Тимофеев-Ресовский.
ГЕНОКОПИЯ, миметические гены, возникновение сходных фенотипич признаков под влиянием генов, расположенных в разных участках хромосомы или в разных хромосомах (т. н. мутантные аллели). Явление Г., установленное прежде всего на высших организмах, свидетельствует о сложном характере наследования мн. признаков. Биохим. природа Г. заключается в наличии в клетке неск. параллельных путей синтеза тех или иных её компонентов (напр., синтез тимидиловой к-ты в бактериальной клетке может осуществляться как из уридиловой, так и из цитидиловой к-т). Разные мутации, действие к-рых реализуется через один и тот же процесс или орган, могут с неодинаковой полнотой копировать друг друга по своему конечному эффекту; в свою очередь, их конечный эффект может имитироваться при действии различных внеш. факторов (см. Фенокопия). Явления Г. и фенокопии очень важны для понимания механизмов реализации наследственных (при Г.) и ненаследственных (при фенокопиях) аномалий и болезней у человека. В.
Н Сойфер, В. П. Эфроимсон.
ГЕНОМ [нем. Genom, англ, genom(e)], гаплоидный хромосомный набор; совокупность генов, локализованных в одиночном наборе хромосом данного организма. Термин предложен в 1920 нем. биологом Г. Винклером. Под Г. принято понимать совокупность генов, сосредоточенных в хромосомах без учёта наследственных детерминант, связанных со структурами цитоплазмы. В гаметах диплоидных организмов, а также в клетках гаплоидных организмов содержится один Г.; в соматич. клетках диплоидных организмов - два Г. С увеличением степени плоидности клеток растёт число Г. При оплодотворении 'происходит объединение Г. отцовских и материнских гамет. Как правило, Г., полученные от отцовской и материнской гамет, гомологичны. Гомология между всеми или неск. Г. отсутствует лишь у отдалённых гибридов. Под абс. гомологией двух Г. понимают совпадение линейного расположения генов в каждой хромосоме. Наличие такого совпадения обеспечивает возможность нормальной конъюгации хромосом в мейозе. Изменения числа хромосом (напр., полиплоидизация, увеличение числа или выпадение отдельных хромосом) наз. геномными мутациями. Организм, у к-рого неск. раз повторяется один и тот же Г., наз. автополиплоидом (см. Автополиплоидия). Организм, в к-ром объединены разные Г., наз. аллополиплоидом (см. Аллополиплоидия). Примером аллополиплоидов могут служить пшеницы. Гаплоидное число хромосом у твёрдой пшеницы - 14, у мягкой - 21, у однозернянки - 7. Путём гибридизации и изучения конъюгации хромосом в мейозе было выяснено, что один Г. из 7 хромосом имеется у всех пшениц (геном А); у твёрдой и мягкой - два общих Г. по 7 хромосом (А, В) и, наконец, у мягкой есть ещё особый геном D, также включающий 7 хромосом. Т. о., геномная формула однозернянки - АА, твёрдой - ААВВ, мягкой пшеницы, возникшей в процессе эволюции путём скрещивания трёх разных диких злаков и удвоения числа хромосом у гибридов,- AABBDD. В опытах сов. генетика Г. Д. Карпеченко были впервые совмещены в гибридном организме Г. редьки и капусты. Путём соответств. скрещиваний и цитологич. анализа можно установить происхождение отдельных Г. Напр., В. А. Рыбин от скрещивания алычи и тёрна получил (ресинтезировал) культурную сливу; т. о. было установлено, что Г. сливы включает Г. алычи и тёрна (см. Амфидиплоиды, Геномный анализ). Для понимания структуры и функционирования Г. большое значение имело установление строения молекул нуклеиновых кислот (ДНК и РНК), механизмов их репликации, способов записи и передачи генетич. информации (см. Генетический код). В.
Н. Сойфер, В. В. Хвостова.
ГЕНОМНЫЙ
АНАЛИЗ, анализ происхождения
различных геномов у полиплоидных форм посредством скрещиваний между
предполагаемыми родительскими формами с последующей полиплоидизацией (см.
Плоидность, Полиплоидия). Один из цитогенетич. методов; предложен
япон. цитогенетиком X. Кихарой (1924). При подборе пар для скрещивания
ориентируются на морфологич. признаки, общие для обоих партнёров и изучаемой
аллополишюидной формы (см. Аллополиплоидия). После скрещивания и
полиплоидизации проводят точный хромосомный анализ и анализ процесса мейоза.
Конъюгация хромосом служит показателем родства геномов. Отсутствие
конъюгации свидетельствует о принадлежности хромосом к разным геномам.
ГЕНОНЕМА (от
ген и греч. пета - нить), длинная белковая молекула-нить (или их
пучок), к-рая, согласно модели, предложенной в 1928 сов. биологом Н. К.
Кольцовым, представляет основу хромосомы и является носителем генетич.
информации. Радикалы Г. рассматривались как гены, а атомные изменения в
белковой молекуле - как причины мутаций. Эта модель после открытия роли
дезоксирибонуклеиновой кислоты устарела, но выдвинутое Кольцовым
при её построении предположение о способности хромосомы к самокопированию
было подтверждено дальнейшими исследованиями.
ГЕНОТИП (от ген и тип), совокупность всех генов, локализованных в хромосомах данного организма. В более широком смысле Г.- совокупность всех наследственных факторов организма-как ядерных (геном), так и неядерных, внехромосомных (т. е. цитоплазматических и пластидных наследственных факторов). Термин предложен датским биологом В. Иогансеном (1909). Г.- носитель наследственной информации, передаваемой от поколения к поколению. Он представляет собой систему, контролирующую развитие, строение и жизнедеятельность организма, т. е. совокупность всех признаков организма- его фенотип. Г.- единая система взаимодействующих генов, так что проявление каждого гена зависит от генотипической среды, в к-рой он находится. Напр., красная окраска цветков у нек-рых сортов душистого горошка возникает только при одновременном присутствии в Г. доминантных аллелей двух различных генов, тогда как порознь каждая из этих аллелей обусловливает белую окраску цветков (см. Комплементация). Взаимодействие Г. с комплексом факторов внутр. и внеш. среды организма обусловливает фенотипическое проявление признаков. Примером влияния среды на фенотипич. проявление Г. может служить окраска меха у кроликов т. н. гималайской линии: при одном и том же Г. эти кролики при выращивании на холоде имеют чёрный мех, при умеренной темп-ре - гималайскую окраску (белая с чёрными мордой, ушами, лапами и хвостом), при повыш. темп-ре - белый мех. Потомки этих трёх групп животных наследуют не какую-то одну неизменную окраску меха, а способность давать определ. окраску, различную в разных условиях среды. Поэтому в общем виде правильнее говорить, что Г. определяет наследование не конкретных признаков, а норму реакции организма на все возможные условия среды. На разных этапах развития особи в активном состоянии находятся то одни, то др. гены; поэтому Г. в онтогенезе функционирует как изменчивая подвижная система. Термин Г. иногда употребляют в более узком смысле для обозначения лишь группы генов или даже отдельных генов, наследование к-рых составляет предмет наблюдения. Напр., в расщепляющемся потомстве от моногибридного скрещивания АА X аа принято говорить о генотипах АА, Аа и аа, отвлекаясь от возможных различий между соответствующими особями (или группами особей)по др. генам. В.И.Иванов.
ГЕНОТИПИЧЕСКАЯ
СРЕДА, генетический фон,
совокупность генов, оказывающих влияние на проявление в фенотипе (структурах
и функциях организма) данного гена. Термин предложен сов. генетиком
С. С. Четвериковым в 1926. В дальнейшем экспериментально установлено, что
проявление каждого гена зависит от влияния др. генов. Это, в частности,
доказано существованием генов-регуляторов. С углублением исследований клетки
как целостной системы выясняется всё большее значение роли Г. с., прежде
всего во взаимосвязанных процессах обмена веществ.
ГЕНОФОНД (от ген и фонд), качественный состав и относительная численность разных форм (аллелей) различных генов в популяциях и населениях того или иного вида организмов. Термином Г. (введён рус. учёным А. С. Серебровским в 1928) обозначают аллельный состав популяции или всего населения вида, включая все варьирующие признаки и свойства вида или же ту или иную интересующую исследователя выборку из них. Рецессивный Г.- в основном укрытая от естеств. отбора совокупность рецессивных аллелей-при резком сокращении численности особей и изменившихся внеш. условиях обеспечивает быструю наследственную перестройку популяции. У генетически менее изученных видов можно определять т. и. фенофонд (фен - элементарный признак). Инвентаризация наследственно варьирующих признаков изучаемого вида с.-х. животных и растений совместно с определением частот различных аллелей имеет большое практич. значение. Изучение Г. человека имеет фундаментальное значение для генетики человека. Лит.: СеребровскийА. С., Геногеография и генофонд сельскохозяйственных животных СССР, Научное слово, 1928, Jsfe 9. Н.
В. Тимофеев-Ресовский.
ГЕНОЦИД (от греч. genos - род, племя и лат. caedo - убиваю), истребление отдельных групп населения по расовым, национальным или религиозным мотивам, одно из тягчайших преступлений против человечества. Преступления Г. органически связаны с фашизмом и аналогичными реакционными теориями, пропагандирующими расовую и национальную ненависть и нетерпимость, господство т. н. высших рас над низшими и т. п. Преступления Г. совершались в массовых масштабах гитлеровцами во время 2-й мировой войны 1939-45 в оккупированных странах Европы, особенно против славянского и еврейского населения. Миллионы людей различных национальностей были уничтожены в фашистских лагерях смерти. Правящие круги ряда империалистич. гос-в грубо нарушают Конвенции 1948 и 1965, проводя политику Г. как внутри страны, так и на подвластных им территориях, используя эту политику для борьбы против национально-освободительного движения. Политика Г. и апартхейда стала гос. политикой в Южно-Африканской Республике, в Родезии. Наказуемость Г. установлена уставами междунар. военных трибуналов (Нюрнбергского и Токийского), а также спец. междунар. конвенцией О предупреждении преступления геноцида и наказании за него (одобрена Генеральной Ассамблеей ООН 9 дек. 1948). Согласно конвенции, под Г. понимаются действия, совершаемые с намерением уничтожить, полностью или частично, к.-л. национальную, этническую, расовую или религиозную группу как таковую, а именно: убийство членов такой группы, причинение им серьёзных телесных повреждений или умственного расстройства; предумышленное создание условий, к-рые рассчитаны на полное или частичное физич. уничтожение таких групп, принятие мер, рассчитанных на предотвращение деторождения в их среде, насильственная передача детей из одной человеческой группы в другую. Конвенция предусматривает также наказуемость заговора с целью совершения Г., подстрекательства, покушения и соучастия в Г. Конвенция предусматривает предупреждение ц наказание преступлений физич. и бпол. Г. При выработке конвенции представитель СССР настаивал на запрещении также национально-культурного Г., к-рый выражается в мероприятиях и действиях, направленных против пользования национальным языком и против национальной культуры к.-л. группы населения, однако империалистич. державы отказались принять это предложение, а также распространить действие конвенции на колонии, в к-рых преступления Г. носят массовый характер. В 1965 ООН принята конвенция о ликвидации всех форм расовой дискриминации, осуждающая расизм, одной из форм которого является Г. В Советском
Союзе и других социалистич. гос-вах всякое ограничение прав или установление
к.-л. привилегий граждан в зависимости от их расовой, национальной или
религиозной принадлежности, а также проповедь расовой или нац. исключительности
или ненависти и пренебрежения запрещены и караются законом.
ГЕНРИ (Henry) Джозеф (17.12.1797, Олбани, - 13.5.1878, Вашингтон), американский физик. С 1832 проф. Прин-стонского колледжа, с 1846 секретарь и директор Смитсоновского ин-та, с 1868 президент Нац. АН США; первый президент Философского об-ва в Вашингтоне (с 1871). Осн. труды по электротехнике. В 1828 впервые построил электромагниты большой силы, применив многослойные обмотки из изолированной проволоки. Г. открыл явление самоиндукции (1832) и колебательный характер разряда конденсатора (1842). Его именем названа единица индуктивности в системе СИ - генри. Г. принадлежат также работы по метеорологии. Соч.: Scientific writings, v. 1 - 2, Wash., 1886. Лит.: Лебедев
В. И., Изобретение Джозефа Генри, Вестник связи, 1946, № 8.
ГЕНРИ (О.
Henry) (1862-1910), американский писатель; см. О. Генри.
ГЕНРИ (Henry)
Уильям (12.12.1774, Манчестер,-2.9.1836, Пендлбери), английский химик.
Доктор медицины Эдинбургского ун-та (1807). Открыл зависимость растворимости
газов в воде от температуры и давления (см. Генри закон).
ГЕНРИ, единица индуктивности и взаимной индуктивности в Международной системе единиц и МКСА системе единиц. Названа в честь амер. учёного Дж. Генри. Сокращённое обозначение: русское гн, междунар. Н. 1 генри равен индуктивности электрич. контура, возбуждающего магнитный поток в 1 вебер при силе постоянного тока в нём 1 ампер. Г. может быть также определён как индуктивность электрич. цепи, в к-рой возникает эдс самоиндукции в 1 вольт при равномерном изменении тока в этой цепи со скоростью 1 ампер в 1 сек. На практике часто пользуются дольными единицами: миллигенри (10-3 гн) и микрогснри (10-6 гн). Генри на метр - единица абсолютной магнитной проницаемости, равная абс. магнитной проницаемости среды, в к-рой при напряжённости магнитного поля 1 а/м создаётся магнитная индукция 1 тл. Г.
Д. Бурдун.
ГЕНРИ ЗАКОН, положение, согласно к-рому при постоянной темп-ре растворимость газа в данной жидкости (выраженная весовой концентрацией его) прямо пропорциональна давлению этого газа над раствором. Г. з. описан У. Генри в 1803; он хорошо соблюдается только для идеальных растворов и применим лишь в области невысоких давлений, приобретая при этом характер предельного закона. О растворимости газовых смесей см. Дальтона законы. В.
А. Киреев.
ГЕНРИЕТТЫ
ОСТРОВ, один из о-вов архипелага
Де-Лонга в Восточно-Сибирском м. Пл. 12 км2. Сложен гл.
обр. песчаниками. Вые. 315 м. Покрыт ледниками.
ГЕНРИХ (Henry). В Англии:Г. I [1068, Селби(?),-1.12.1135, Лион-ла-Форе], король с 1100. Младший сын короля Вильгельма I Завоевателя. Незаконно захватил власть, склонив на свою сторону англ, баронов обещанием гарантировать соблюдение их феод, прав и привилегий. Опираясь на мелкое рыцарство, растущие города и свободное крестьянство, Г. I значительно укрепил гос. аппарат. При нём оформился центр, орган управления - королевская курия. Г. I вступил в конфликт с римским папой по вопросу о праве назначения англ, епископов. По соглашению 1107 король сохранил право на получение феод, присяги от англ, прелатов при их вступлении во владение церк. землями. Г. II (5.3.1133, Ле-Ман,-6.7.1189, Шинон), король с 1154, первый из династии Плантагенетов. Под властью Г. II наряду с англ, владениями находились обширные владения во Франции. Став королём после долгой феодальной смуты (1135-53), Г. II восстановил мир и добился роспуска баронских наёмных отрядов. Опираясь на рыцарство, горожан и верхушку свободного крестьянства, проводил политику дальнейшего укрепления централизованного феод, гос-ва в Англии. Судебная реформа Г. II значительно расширила компетенцию королев, судов за счёт феод, курий и ввела в них расследование через присяжных; отныне любой свободный мог за плату обратиться в королев, суд. Попытка Г. II с помощью т. н. Кларендонских постановлений (1164) подчинить королев, власти церк. суды не увенчалась успехом. В результате воен. реформы широкое распространение получила замена рыцарской службы ден. платежом, т. н. щитовыми деньгами; кроме того, все свободные были обязаны иметь оружие, чтобы при необходимости служить в войске короля. При Г. II были заложены основы всей суд.-адм. системы англ. феод, гос-ва: королев, курия разделилась на высший королев, суд и казначейство, начало складываться англ, общегос. феод, право. При всей прогрессивности реформ Г. II они носили чисто феод, характер. Проводя резкую грань между свободными и крепостными, они юридически закрепляли бесправие последних. При Г. II в 1169 началось завоевание Ирландии. Лит.: Boussard J., Le gouvernement d'Henry II, Plantagenet, P., 1956. E. В. Гутнова. Г. III (1.10.1207, Уинчестер,-16.11. 1272, Лондон), король (с 1216) из династии Плантагенетов, сын Иоанна Безземельного. Пытался управлять страной, опираясь на иностр. феодалов-авантюристов и союз с рим. курией. Недовольство баронов этой политикой в кон. 50-х гг. нашло поддержку у мелкого рыцарства, горожан и зажиточного крестьянства. В 1258 Г. III был вынужден утвердить Оксфордские провизии, установившие в стране режим баронской олигархии, а затем Вестминстерские провизии, неск. ограничивавшие произвол баронов. Отказ Г. III от соблюдения •Оксфордских провизии привёл к гражд. войне (1263-67). Симон де Монфор в 1264 разбил королевские войска и взял в плен Г. III. После разгрома сторонниками короля войск оппозиции и гибели Монфора (авг. 1265) Г. III в 1266 был полностью восстановлен в своих правах. Однако возросшая роль рыцарства и городов вынудила Г. III и баронов установить практику созыва парламента. Лит.: Гутнова Е. В., Возникновение английского парламента, М., 1960. Е. В. Гутнова. Г. IV [апр. 1366(?), замок Болингброк, графство Линкольншир, - 20.3.1413, Лондон], король с 1399, основатель династии Ланкастеров. В 1397 поддержал гос. переворот Ричарда II, но вскоре был изгнан. В 1399 высадился в Англии и возглавил мятеж магнатов Сев. Англии против Ричарда II. После отречения последнего был провозглашён королём. Правление Г. IV прошло в борьбе с мятежами крупных феодалов, восстаниями в Уэльсе, набегами шотландцев. Г. IV искусно разъединял своих противников, стремился опираться на мелкое дворянство и зажиточных горожан и заручиться поддержкой парламента, права к-рого были значительно расширены. Жестоко подавлял демократич. движения; в 1401 провёл статут, направленный против лоллардов. Ю. Р. Ульянов. Г. V[16.9(?). 1387, Монмут,-31.8.1422, Буа-де-Венсенн], король (с 1413) из династии Ланкастеров, сын Генриха IV. Стремился примириться с феод, знатью, возвратив ей владения, конфискованные Генрихом IV. В 1414 подавил восстание лоллардов. Возобновив Столетнюю войну, нанёс французам в 1415 решит, поражение при Азенкуре и вскоре захватил С. Франции с Парижем. Был признан наследником франц. короля Карла VI и регентом Франции. Ю. Р. Ульянов. Г. VI (6. 12.1421, Виндзор,-21.5.1471, Лондон), король (1422-61) из династииЛанкастеров, сын Генриха V. Вступил на престол 9-месячным ребёнком. Слабовольный и подверженный всё более продолжительным припадкам безумия, был игрушкой в руках часто сменявшихся регентов и фаворитов. В 1461, в ходе Алой и Белой розы войны, был низложен королём Эдуардом IV Йорком. После врем, реставрации (3.10.1470- 11.4.1471) Г. VI был вторично низложен и убит в Тауэре. Г. VII (28.1.1457, Пембрук,-21.4. 1509, Ричмонд), король с 1485, основатель династии Тюдоров. Вступил на престол как кандидат Ланкастеров после победы над Ричардом III в битве при Босворте (22 авг. 1485), знаменовавшей завершение войны Алой и Белой розы. Провёл ряд мероприятий, ограничивавших могущество феод, знати [конфискация зем. владений, роспуск воен. дружин, создание чрезвычайных судов для расследования дел о заговорах (Звёздная палата, с 1487)]. В интересах нарождавшейся буржуазии способствовал развитию судоходства и внешней торговли. В правление Г. VII были заложены основы абсолютизма. В. Ф. Семёнов. Г. VIII (28.6.1491, Гринвич,-28.1. 1547, Лондон), король с 1509, 2-й из династии Тюдоров; один из ярких представителей англ, абсолютизма. В своём правлении опирался на узкий круг фаворитов (Т. Уолси, а позднее Т. Кромвель и Т. Кранмер). В правление Г. VIII была проведена Реформация, к-рую он рассматривал как важное средство укрепления абсолютизма и королев, казны. Поводом к Реформации послужил отказ папы утвердить развод Г. VIII с Екатериной Арагонской и его женитьбу на Анне Болейн. В 1534 после разрыва с папой был провозглашён парламентом главой английской (англиканской)церкви, сохранившей католич. обряды. Т. Мор (лорд-канцлер с 1529), выступивший против Реформации, был казнён (1535). В 1536 и 1539 была проведена секуляризация монастырей, земли к-рых перешли в руки нового дворянства. Сопротивление этой политике, особенно на С., жестоко подавлялось (см. Благодатное паломничество). Громадные расходы двора, войны с Францией и Шотландией привели в конце царствования Г. VIII к полному расстройству финансов. В связи с усилившейся в результате секуляризации экспроприацией крестьянства издал статуты против бродяг и нищих (см. Кровавое законодательство против экспроприированных). Хотя политика Г. VIII отвечала в нек-рой степени интересам нового дворянства и растущей буржуазии, его классовой опорой было феод, дворянство (попытки Г. VIII сохранить старую феод, структуру землевладения в эпоху начавшегося агр. переворота отразились, в частности, в его мероприятиях по ограничению огораживаний). Лит.: Масkie J. D.. The earlier Tudors. 1485-1558, Oxf., 1952; Scar i sbrick J. J., Henry VIII,L., 1969. В. Ф. Семёнов.
ГЕНРИХ (Heinrich). В королевстве Германия и Священной Римской империи. Наиболее значительны: Г. I (ок. 876-2.7.936, Мемлебен, Саксония), король с 919, основатель Саксонской династии. Саксонский герцог из рода Людольфингов. От вынужденной политики уступок усилившимся герцогам Г. I постепенно переходил к их подчинению. Вновь присоединил (925) к Герм, королевству отпавшую ранее Лотарингию. Для борьбы с набегами венгров выстроил и укрепил ряд бургов и создал сильную конницу; одержал победу над венграми при Риаде на р. Унструт (933). Воен. походами 928- 929 начал захват земель полабских славян. Политика Г. I подготовила значит, усиление королев, власти при его сыне и преемнике Оттоне I. Лит.: Bartmuss H. J.. Die Geburt des ersten deutschen Staates, В., 1966. Г. III (28.10.1017-5.10.1056), король с 1039, император с 1046, из Фоанконской династии; сын Конрада II. Опирался на министериалов и рыцарство. Во время похода в Италию (1046-47) низложил соперничавших пап, после чего неоднократно назначал кандидатов на папский престол. Однако покровительство Г. III церк. клюнийской реформе подготовило почву для последующего усиления папской власти. Поставил Чехию и Венгрию в зависимость от империи; вынудил к подчинению герцога Лотарингского. Г. IV (11.11.1050-7.8.1106, Льеж), король с 1056, император с 1084, сын Генриха III. В период малолетства Г. IV в Германии усилились князья, расхищавшие владения короны. Мероприятия Г. IV по укреплению позиций королев, власти в Саксонии вызвали Саксонское восстание 1073-75, с трудом им подавленное. Стремление сохранить за собой право церк. инвеституры в Геомании и Сев. Италии привело Г. IV (в 1076) к столкновению с папой Григорием VII; разговелась длит, борьба за инвеституру. Отлучённый папой от церкви и низложенный, Г. IV был вынужден под давлением князей пойти на покаяние к папе в Каноссу (янв. 1077). Вновь отлучённый от церкви (1080), в 1084 овладел Римом и был коронован своим ставленником антипапой Климентом III, но отступил перед союзниками Григория VII - норманнами Юж. Италии. Объединение пап с нем. князьями, неудачный 3-й поход в Италию (1090-97), восстания против Г. IV его сыновей и его пленение не заставили изворотливого и энергичного Г. IV признать себя побеждённым. Ему удалось бежать из плена, но во время подготовки новой войны с сыном он умер. В целом попытка Г. IV упрочить королев, власть (путём укрепления королев, домена, опоры на министеоиалитет и отчасти на города) окончилась полным крахом в условиях роста раздробленности Германии. Лит.: SchmeidlerR., Kaiser Heinrich IV, Lpz., 1927; Stern L. und Gericke H.. Deutschland von der Feudalepoche von der Mitte des 11. Jahrhunderts bis zur Mit-te des 13. Jahrhunderts, В., 1964. М. Л. Абрамсон.
Г.
V (8.1.1081-23.5.1125, Утрехт), король с 1106, император с 1111, сын
Генриха IV. При жизни отца сблизился с его противниками - папой и нем.
князьями. После смерти Генриха IV возобновил борьбу с папством за инвеституру,
окончившуюся компромиссным Вормсским конкордатом (1122). Со смертью
Г. V прекратилась Франконская династия.
F.VI (1165,
Нимвеген,-28.9.1197, Мессина), король с 1190, император с 1191 из династии
Штауфенов. Сын Фридриха I Барбароссы. Благодаря женитьбе на наследнице
сицилийского короля Констанции (1186) присоединил к владениям Штауфенов
Сицилийское королевство (утвердился там лишь в 1194).
Г.
VII (ок. 1275-24.8.1313, Буонконвенто, близ Сиены), король с 1308,
император с 1312, первый из династии Люксембургов. Добился в 1310 передачи
своему сыну Иоанну чеш. престола. В 1310 вторгся в Италию, безуспешно пытаясь
силой вновь подчинить её империи.
ГЕНРИХ (Henri).
Во Франции. Наиболее значительны:
Г.
II (31.3.1519, Сен-Жермен-ан-Ле,- 10.7.1559, Париж), король с 1547.
Из династии Валуа. Был женат (с 1533) на Екатерине Медичи. Находился
под влиянием коннетабля Анн де Монморанси, особенно в области внешней
политики. Жестоко преследовал гугенотов, учредив для суда над ними в 1547
Огненную палату, в 1559 издал эдикт, требовавший для еретиков смертной
казни. В союзе с немецкими протестантскими князьями вёл борьбу против имп.
Карла V; в 1552 занял епископства Мец, Туль, Верден. В 1558 отвоевал у
англичан Кале. В 1559 подписал Като-Камбрезийский мир, завершивший Итальянские
войны. Был смертельно ранен на турнире.
Лит.: Noell
H., Henri П..., Р.,
1944.
Г.
III (19.9.1551, Фонтенбло,-2.8.1589, Сен-Клу), король с 1574, последний
представитель династии Валуа. Был избран в 1573 на польск. престол,
но, узнав о смерти своего брата франц. короля Карла IX, тайно бежал из
Польши, чтобы занять франц. престол. Г. III правил в разгар Религиозных
войн. Боролся как с гугенотами, к-рых возглавлял Генрих Наваррский,
так и с Гизами, вождями Католической лиги, претендовавшими на наследование
престола в связи с бездетностью Г. III. В мае 1588 в день баррикад (восстание
в Париже, организованное Парижской лигой - объединением, игравшим
решающую роль в Католич. лиге 1585-96) Г. III бежал в Шартр. В том же году
после убийства герцога Гиза и его брата кардинала Лотарингского, совершённого
по приказу Г. III, демократич. крыло Парижской лиги низложило Г. III. Последний
пошёл на соглашение с Генрихом Наваррским, и они совместно осадили Париж.
При осаде Г. III был убит подосланным лигой монахом Жаком Клеманом.
Соч.:
Lettres d'Henri III, t. 1 - 2, P., 1959-65.
Лит.: Erlanger
Ph., Henri III, P., [1935]: Champion P., Henri III, [v.1-2], P., [1943
- 51]. А. Д. Люблинская.
Г. IV
(13. 12. 1553, По, Беарн,- 14.5.1610, Париж), король с 1589 (фактически
с 1594), первый из династии Бурбо-нов; король Наварры (Генрих Наваррский)
с 1562. Во время Религиозных войн - глава гугенотов. В 1593
принял католичество и вступил в Париж. Нантским эдиктом 1598 предоставил
гугенотам свободу вероисповедания и многие привилегии. Политика Г. IV способствовала
укреплению абсолютизма. Ген. штаты перестали созываться; компетентность
провинц. собраний была ограничена; возросла роль бюрократич. аппарата,
чиновники добились официального закрепления права наследования и продажи
должностей (см. Полетта). Г. IV отнял у губернаторов значит, долю
гражданской власти, усилив полномочия ген. наместников и начинавших играть
известную роль интендантов провинций. В области экономики проводил
политику протекционизма, поощрял развитие франц. мануфактур; ввёл покровительств.
таможенный тариф (1599); при нём проводились работы по улучшению дорог,
строительству каналов и т. п. В 1604 началась колонизация французами Канады.
Правительством Г. IV были отменены недоимки с крестьян за прошлые годы,
снижена талья, но одновременно были увеличены косвенные налоги (на
соль, вино и др.). Г. IV играл гл. роль в организации антигабсбургской
оппозиции, вёл подготовку к войне с Габсбургами. Был убит католиком-фанатиком
Равальяком.
Генрих
IV.
Гравюра
X. Голциуса.
Лит.: М
осина 3., Абсолютизм в политике Генриха IV, Историк-марксист, 1938, кн.
2:Пыхтеев Б., Мероприятия Генриха IV по развитию сельского хозяйства во
Франции. Уч. зап. Московского государственного педагогического ин-та. 1940, т.
26; De Vaissiere P., Henri IV, P., [1928]; Ritter R., Henri IV, luimeme. L'Homme,
P., [1944]. А. Д. Люблинская.
ГЕНРИХ
ЛАТВИЙСКИЙ (Heinrich von
Lettland) (1187-1259), автор Хроники Ливонии (см. Ливонские хроники),
написанной в 1224-27 на латинском яз. Онемеченный католич. священник.
Прибыл в Ригу в 1205. С 1208 священник латышского прихода в Паппендорфе.
Участник походов нем. крестоносцев в Ливонию. Восхвалял захватнич. политику
рыцарей. В Хронике подробно описывается агрессия нем. феодалов в Ливонии
и Эстонии с кон. 12 в. по 1227.
Соч.:
Heinrici Chronicon Livoniac. 2
Aufl., Hannoverae, 1955; в рус. пер.- Хроника Ливонии, М. -Л., 1938. Х.Х.Круус.
ГЕНРИХ
ЛЕВ (Heinrich der Lowe)
(1129-6.8.1195, Брауншвейг), герцог Саксонии (в 1142-80) и Баварии (в 1156-80)
из рода Вельфов. Вместе с Альбрехтом Медведем возглавил крестовый
поход против славян 1147, окончившийся неудачей. В результате последующих
походов (с 1160) захватил почти всю терр. бодричей и стал владельцем
огромной терр. к В. от Эльбы. Истребляя и оттесняя славян к востоку, Г.
Л. переселял на захваченные земли нем. колонистов. Усиление Г. Л. вызвало
резкий конфликт между ним и имп. Фридрихом I Барбароссой. Последний, используя
отказ Г. Л. от участия в походе императора в Италию, организовал суд. процесс
над Г. Л. (1180). Г. Л. был лишён большинства владений (в его руках остались
только Брауншвейг и Люнебург).
ГЕНРИХ
МОРЕПЛАВАТЕЛЬ (Dom Enrique
о Nayegador) (4.3. 1394, Порту,- 13.11.1460, Сагрищ), португальский принц,
организатор морских экспедиций к о-вам центр, части Атлантич. ок. и берегам
Африки (за что в 19 в. получил прозвище Мореплаватель, хотя сам не плавал).
Г. М. на средства Ордена Христа (главой к-рого он стал) основал в Сагрише
(Португалия) обсерваторию и мореходную школу, способствовал развитию португ.
кораблестроения (преобладающей стала каравелла). Океанские экспедиции Г.
В. Кабрала, А. Кадамосто и др. открыли о-ва Азорские (1432-35), Зелёный
мыс, pp. Сенегал и Гамбия, о-ва Бижагош (1434-57) и о-ва Зелёного Мыса
(1456). По инициативе Г. М. начался вывоз афр. рабов в Португалию; капитаны
Н. Триштан, Д. Диаш, А. Фернандиш и др., в поисках золота и охотясь за
рабами, в 1434-60 обследовали и нанесли на карту ок. 3500 км зап.-афр.
побережья от Зап. Сахары до Гвинейского зал. Экспедиции Г. М. положили
начало португ. экспансии в Африку.
Лит.: Магидович
И. П., Очерки по истории географических открытий, М., 1967: S a n с е a
u E., Henry, the Navigator..., N. Y., 1947.
ГЕНРИХ
НАВАРРСКИЙ, король Наварры
с 1562, франц. король Генрих IV.
ГЕНСЛЕ (польск.
gesle), 1) общее наименование старинных польских струнных инструментов.
2) Г. подгалянские, или злубцоки (ziobcoki), старинный польский народный
смычковый инструмент. Корпус деревянный, долблёный грушевидный (как у болг.
гадулки) или клеёный лодкообразный, струн 3-4, строй квинтовый.
При игре инструмент опирали о левую сторону груди ниже плеча. Звук Г.-
резкий, пронзительный.
ГЕНТ, Ган
(флам. Gent, франц. Gand), город в Бельгии, на р. Шельда при впадении в
неё р. Лис, каналами связан с портами Остенде и Тернёзен на Северном м.
Адм. ц. провинции Вост. Фландрия. 229,7 тыс. жит. (1969, с пригородами).
Первое
упоминание о Г. в источниках относится к 7 в. С 11-12 вв. Г. один из центров
суконного произ-ва европ. значения. Непосредственно подчинённый власти
фландрских графов, Г. в 12-13 вв. добился значит, самостоятельности в делах
внутр. гор. управления и стал одним из гл. центров социально-политич. борьбы
во Фландрии. Население Г. участвовало во Фландрском восстании 1323-28,
здесь происходило восстание, возглавленное с 1338 Я. Артевелде, Гентское
восстание 1539-40. Г. - один из центров революц. движения во время
Нидерл. бурж. революции 16 в. В 16 в. в связи с общим упадком цехового
ремесла Г. потерял значение одного из важнейших экономич. центров Европы;
в 19 в. стал вновь экономически возрождаться.
Г.- крупный
пром. центр и трансп. узел. Второй по значению порт и гл. текст, центр
страны. Преим. хл.-бум. и льняное произ-во. Металлургия, текст, и электро-технич.
машиностроение, нефтепереработка, хим., бум., пищ. (муком., пивовар. и
др.) пром-сть. Старинное произ-во кружев. Ввоз хлопка, льна, угля, нефти
и ДР-Г.-центр фламандской культуры. Ун-т (с 1817). Музеи: археологич.,
изящных иск-в.
Гент.
Площадь Синт-Бафсплейн. На переднем плане - городская башня (1183- 1339)
с суконными рядами (1426 - 41,), за ними церковь Синт-Никласкерк (13- 17
вв.).
Ср.-век.
планировка, обилие архит. памятников начиная с романской эпохи (замок графов
Фландрских, 1180- 1200,- овальный в плане, с мощными стенами и башнями;
хлебный склад, ок. 1200), старинная застройка площадей и набережных придают
центру Г. вид города-музея. Особенно богат Г. готич. сооружениями (собор
св. Бавона, ок. 1228-1559, с Гентским алтарём братьев ван Эйк; городская
башня вые. 91 м, 1183-1339; ратуша, 1518-35, арх. Д. де Вагемакере,
Р. Келдерманс; Дом свободных корабельщиков, 1530-31; укреплённый мост Работ,
1489; много-числ. церкви, монастырские постройки, жилые дома). Среди позднейших
сооружений - Дворец юстиции (1836-46, арх. Л. Руландт), библиотека ун-та
(1935-40, арх. X. ван де Велде; илл. см. т. 3, вклейка кс. 185), новые
пром. р-ны.
Лит.: Пиренн
А., Средневековые города Бельгии, пер. с франц., М., 1937; D'Hоndt J.,
Keyser p. d e, Gent, [Antw., 1947]; Fris V.,Histoire de Gand, Brux., 1913;
Keyser P. de, 1000-jarig Gent, Gent, 1949.
ГЕНТЕКС (англ.
Gentex - Generalized Teletype Exchange Service), международная телеграфная
сеть общего пользования, оборудованная автоматич. коммутационными телеграфными
станциями прямых соединений. Входящая в эту сеть телеграфная станция
Г. в Москве соединена международными каналами связи с аналогичными станциями
ряда европейских социалистич. стран. Др. телеграфные станции в Сов. Союзе
получают соединение со станциями Г. набором их номеров через моек, станцию.
ГЕНТ-ОСТЕНДЕ
КАНАЛ, в Бельгии, соединяет
р. Шельда у г. Гент с Северным м. у порта Остенде. Состоит из двух частей:
канала Гент
- Брюгге
(прорыт в 1751) длиной в 47 км и канала Брюгге - Остенде (1622)
длиной в 20,5 км. Глубина 3,1 м. Осн. грузы: уголь, стройматериалы,
с.-х. продукция, хим. изделия, текст, сырьё.
ГЕНТСКИЙ
ДОГОВОР 1814, договор между
Англией и США, завершивший англо-американскую- войну 1812-14. Подписан
24 дек. 1814 в Генте. Предусматривал взаимное возвращение захваченных территорий,
прекращение воен. действий против индейцев, принятие решит, мер к прекращению
работорговли. Восстанавливая старые границы, договор не разрешил спорных
вопросов, вызвавших войну. За Г. д. последовало заключение торг, конвенции
1814, соглашения о демилитаризации Великих озёр 1817, конвенции 1818 о
рыболовстве в Сев. Атлантике, о сев. границе США и т. н. совместном владении
Орегоном.
ГЕНТСКОЕ
ВОССТАНИЕ 1539-40, было непосредственно
вызвано попыткой исп. пр-ва Карла V насильственно собрать налог, на к-рый
Гент не дал согласия. Но причины восстания лежали глубже (кризис
цехового сукноделия, тяжёлые для трудящихся масс последствия процесса первоначального
накопления капитала). Начало открытому восстанию положил захват власти
в городе цехами (19 авг. 1539), его кульминацией был период 2 сент.- 3
нояб., характеризующийся политич. преобладанием гор. плебса и окрестной
бедноты (крезеры). Восстание распространилось и на соседние города. Восставшие
казнили, арестовали или изгнали наиболее ненавистных членов гор. олигархии,
конфисковали их имущество. После попытки (хотя и неудавшейся) патрициата
и богатого бюргерства организовать переворот восстание пошло на убыль.
14 февр. 1540 Карл V с войском вступил в Гент. Участники восстания подверглись
репрессиям. Гент был обложен контрибуцией, лишён всех вольностей.
Лит.: Чистозвонов
А. Н., Гентское восстание 1539-1540 гг., М., 1957.
А.
Н. Чистозвонов.
ГЕНТСКОЕ
УМИРОТВОРЕНИЕ, соглашение,
заключённое в г. Гент 8 нояб. 1576 в ходе Нидерландской буржуазной
революции 16 века между сев. провинциями Нидерландов, восставшими в
1572, и юж. провинциями, на к-рые восстание распространилось в сент. 1576.
Предусматривало совместную борьбу против исп. войск в Нидерландах, амнистию
участникам восстания, сохранение католицизма на Ю. и кальвинизма на С.
страны и др. Владения католич. церкви и власть Филиппа II Испанского формально
оставались неприкосновенными. С созданием революц. Утрехтской унии (1579)
Г. у. фактически утратило силу.
ГЕНТСМАН, правильнее
Хантсмен (Huntsman) Бенджамин (4. 6. 1704, Линкольншир, - 1776, Аттерклифф),
автор тигельного способа получения литой стали (около 1740). Родился в
Англии в немецкой семье. Тигельный процесс был известен, по-видимому, ещё
в древности в странах Азии (Индия и др.), но технология его держалась в
секрете, в Европе её не знали. Г. переплавлял куски сварочного железа и
чугуна (выплавленного на древесном угле) в присутствии нек-рого количества
флюса. Долгое время тигельный процесс был единственным способом получения
литой стали.
Лит.: Beck
L., Die Geschichte des Eisens in technischer und kulturgeschichtlicher
Beziehung, Abt. 3, Braunschweig, 1897.
ГЕНТ-ТЕРНЁЗЕН
КАНАЛ, в
Бельгии и Нидерландах. Г.- Т. к. соединяет р. Шельда у г. Гент с её устьем
у порта Тернёзен. Прорыт в 1827, реконструирован в 1911. Дл. 34 км (на
терр. Бельгии 19,5 км). Ширина 50 м, глуб. 8,75 м. Осн.
грузы: уголь, руды, строит, камень, химич. продукция, с.-х. продукты, текст,
сырьё.
ГЕНУЭЗСКАЯ
КОНФЕРЕНЦИЯ 1922, международная
конференция по экономическим и финансовым вопросам. Проходила в Генуе (Италия)
с 10 апр. по 19 мая при участии представителей 29 государств - Австрии,
Албании, Бельгии, Болгарии, Великобритании, Венгрии, Германии, Греции,
Дании, Исландии, Испании, Италии, Латвии, Литвы, Люксембурга, Нидерландов,
Норвегии, Польши, Португалии, РСФСР, Румынии, Королевства сербов, хорватов
и словенцев, Финляндии, Франции, Чехословакии, Швейцарии, Швеции, Эстонии,
Японии, а также 5 брит, доминионов - Австралии, Индии, Канады, Новой Зеландии
и Юж.-Афр. Союза. Работой делегации РСФСР руководил В. И. Ленин, назначенный
её председателем. Заместителем председателя делегации был Г. В. Чичерин,
к-рый в Генуе, куда Ленин не выезжал, пользовался всеми правами председателя.
Делегация РСФСР (в её состав входили также Л. Б. Красин, М. М. Литвинов,
В. В. Воровский, Я. Э. Рудзутак, А. А. Иоффе, X. Г. Раковский, Н. И. Нариманов,
Б. Мдивани, А. Бекзадян, А. Г. Шляпников) представляла на Г. к. интересы
не только РСФСР, но и всех др. сов. республик (Азербайджанской, Армянской,
Белорусской, Бухарской, Грузинской,Украинской, Хорезмской ), а также интересы
Дальневосточной Республики. США, отказавшиеся участвовать в работе Г. к.
(нота гос. секретаря Ч. Юза от 8 марта 1922), были представлены на ней
наблюдателем - амер. послом в Италии Р. Чайлдом. Среди представителей капиталистич.
гос-в на Г. к. наиболее активную роль играли Д. Ллойд Джордж, Дж. Н. Керзон
(Великобритания); К. Вирт, В. Ратенау (Германия); Л. Факта (Италия); Ж.
Барту, К. Баррер (Франция). Решение о созыве Г. к. было принято по инициативе
Великобритании на совещании Верх, совета Антанты в Канне (Франция) 6 янв.
1922 (см. Каннская конференция 1922). Офиц. целью Г. к. было изыскание
мер к экономич. восстановлению Центр, и Вост. Европы. Но главным вопросом,
стоявшим на конференции, был, по существу, вопрос об отношениях между Сов.
государством и капиталистич. миром после провала попыток свержения Сов.
власти путём воен. интервенции.
Капиталистич.
страны, в первую очередь Великобритания, в поисках преодоления послевоен.
экономич. трудностей пытались вернуть на мировой рынок Сов. Россию (чтобы,
пользуясь её временной экономической слабостью, широко эксплуатировать
её ресурсы), а также Германию и бывших союзников Германии, потерпевших
поражение в 1-й мировой войне 1914-18. Сов. правительство, заинтересованное
в нормализации экономич. и политич. отношений с капиталистич. гос-вами,
согласилось принять участие в работе Г. к. (8 янв. 1922). На конференции,
однако, ведущую роль играли те представители капиталистич. стран, к-рые
вместо делового обсуждения реальных путей установления экономии, связей
с Сов. государством пытались с помощью дипломатич. нажима добиться от Сов.
правительства экономич. и политич. уступок, ведущих к реставрации капитализма
в России; они рассчитывали заставить Сов. государство признать все долги
царского и Врем, пр-в, вернуть иностр. капиталистам национализированные
Советской властью предприятия или возместить стоимость этих предприятий,
ликвидировать монополию внеш. торговли и т. д. Сов. делегация по указанию
В. И. Ленина отвергла эти требования и, в свою очередь, выдвинула контрпретензии
о возмещении Сов. государству убытков, причинённых иностр. интервенцией
и блокадой (если довоен. и воен. долги России были равны 18,5 млрд. золотых
руб., то убытки Сов. государства в результате иностр. интервенции и блокады
составляли 39 млрд. золотых руб.). Вместе с тем, желая найти почву для
соглашения и восстановления экономич. связей с капиталистич. гос-вами,
сов. делегация на Г. к. 20 апр. 1922 заявила, что Сов. правительство готово
признать довоен. долги и преимущественное право за бывшими собственниками
получать в концессию или аренду ранее принадлежавшее им имущество, при
условии признания Сов. государства де-юре, оказания ему финанс. помощи
и аннулирования воен. долгов и процентов по ним. Огромное значение имело
внесённое сов. делегацией на первом же пленарном заседании Г. к. (10 апр.
1922) предложение о всеобщем разоружении. Однако как вопрос о разоружении,
так и вопросы урегулирования взаимных финанс.-экономич. претензий по вине
капиталистич. гос-в не были разрешены на Г. к. Обсуждение финанс.-экономич.
вопросов было продолжено на Гаагской конференции 1922. В ходе Г.
к. сов. дипломатии, использовавшей противоречия в империалистич. лагере,
удалось прорвать единый фронт империалистич. гос-в, пытавшихся добиться
дипломатич. изоляции Сов. государства, и заключить с Германией Рапалльский
договор 1922 (см. в ст. Советско-германские договоры и соглашения).
Публ.:
Генуэзская конференция 1922. Материалы Генуэзской конференции (Подготовка,
отчеты заседаний, работы комиссий, дипломатическая переписка и пр.), М.,
1922; Документы внешней политики СССР, т. 5, М., 1961.
Лит.: Ленин
В. И., Проект директивы заместителю председателя и всем членам генуэзской
делегации, Поли. собр. соч., 5 изд., т. 44; е г о ж е, О международном
и внутреннем положении Советской Республики. Речь на заседании коммунистической
фракции Всероссийского съезда металлистов 6 марта 1922 г., там же, т. 45;
его же, Политический отчет ЦК РКП(б) 27 марта [на XI съезде РКП(б) 27 марта
- 2 апреля 1922], там же; его лее, Проект постановления ВЦИК по отчёту
делегации на Генуэзской конференции, там же; Любимов Н. Н., Эр л их А.
Н., Генуэзская конференция (Воспоминания участников), М., 1963; Рубинштейн
Н. Л., Внешняя политика Советского государства в 1921 - 1925 гг., М., 1953;
История дипломатии, 2 изд., т. 3, М.,1965, с. 249-304.
И.
И.Минц.
|