БОЛЬШАЯ СОВЕТСКАЯ ЭНЦИКЛОПЕДИЯ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ
|
БОЛЬШАЯ СОВЕТСКАЯ ЭНЦИКЛОПЕДИЯ
В ЭНЦИКЛОПЕДИИ СОДЕРЖИТСЯ БОЛЕЕ 100000 ТЕРМИНОВ |
БИООРГАНИЧЕСКАЯ ХИМИЯ, научное направление, сложившееся на стыке ряда отраслей химии и биологии. Б. х. возникла во 2-й пол. 20 в. и развивается в тесной связи с молекулярной биологией, биохимией и др. биол. дисциплинами, к-рые одновременно исследуют ряд важнейших типов веществ. Б. х. изучает вещества, лежащие в основе процессов жизнедеятельности, в непосредств. связи с познанием их биол. функции. Осн. объекты Б. х.- биополимеры (белки и пептиды, нуклеиновые к-ты и нуклеотиды, липиды, полисахариды и т. д.), превращения к-рых составляют химич. сущность биол. процессов, и биорегуляторы (ферменты, витамины, гормоны, в т. ч. и фи-тогормоны, и др., а также синтетич. биологически активные соединения, напр, лекарств, препараты, ростовые вещества, инсектофунгициды, гербициды и т. д.), к-рые химически регулируют обмен веществ. Б. х. занимается получением этих веществ в химически чистом состоянии, установлением строения, синтезом, выяснением зависимостей между строением и биол. свойствами, изучением хим. аспектов механизма биол. действия биополимеров, а также природных и синтетич. биорегуляторов. Характерная особенность Б. х.- использование всего комплекса хим. и физ. методов индивидуализации веществ (хроматографич. и электро-форетич. методы, противоточное распределение и др.) и выяснение их строения (ультрафиолетовая, инфракрасная, а также Раман-спектроскопия, ядерный магнитный резонанс, электронный и протонный резонанс, химич. масс-спектрометрия, рентгеноструктурный анализ и т. д.). Решение осн. проблем Б. х. важно для дальнейшего прогресса биологии. Без выяснения строения и свойств важнейших биополимеров и биорегуляторов нельзя познать сущность жизненных процессов, а тем более найти пути управления такими сложными явлениями, как размножение и передача наследственных признаков, нормальный и злокачественный рост клеток, иммунитет, память, передача нервного импульса и мн. др. В то же время изучение высокоспециализированных биологически активных веществ и процессов, протекающих с их участием, может открыть принципиально новые возможности для развития химии, хим. технологии и техники. .К проблемам, решение к-рых связано с исследованиями в области Б. х., относятся: создание строго специфичных высокоактивных катализаторов (на основе изучения строения и механизма действия ферментов), прямое превращение хим. энергии в механическую (на основе изучения мышечного сокращения), использование в технике хим. принципов хранения и передачи информации, осуществляемых в биол. системах, принципов саморегулирования многокомпонентных систем клетки в первую очередь избират. проницаемости биол. мембран, и мн. др. Перечисленные проблемы лежат далеко за пределами собственно Б. х.; однако она создает осн. предпосылки для разработки этих проблем, обеспечивая главные опорные пункты для развития биохимич. исследований, относящихся уже к области молекулярной биологии. Широта и важность решаемых проблем, разнообразие методов и тесная связь с др. науч. дисциплинами обеспечили быстрое развитие Б. х. Лит.: Шемякин М. М., Современные проблемы биоорганической химии, М.. 1965; Развитие органической химии в СССР, М., 1967, с. 509 - 73; Хохлов А. С., Овчинник о в Ю. А., Химические регуляторы биологических процессов, М., 1969; Bioorga-nic chemistry, S. F., 1968. А. С. Хохлов. БИООРИЕНТАЦИЯ (от био... и ориентация), способность организмов определять своё местонахождение в пространстве, выбирать оптимальное положение по отношению к действующим на него силам (факторам среды) и определять биологически целесообразное направление движения. Б.- одно из осн. условий приспособления организмов к окружающей среде (адаптации), что может осуществляться тремя путями: изменением состояния организма в соответствии с меняющимися условиями (морфо-физиол. адаптация); сменой мест обитания (см. Кочёвки животных, Миграции животных): изменением обстановки путём образования скоплений (стай, стад и т. п.) или постройки убежищ (нор, гнёзд и т. п.). Б. основана на свойстве раздражимости и восприятия внеш. воздействий физич., химич. и биол. природы. У высших беспозвоночных (членистоногие, моллюски) и у позвоночных животных восприятие, или рецепция, внеш. воздействий (сигналов) осуществляется спец. органами чувств, а их реакции Б. приобретают характер сложных инстинктов, лежащих в основе бионавигации. Выбор направления при передвижениях осуществляется на основании рецепции химич., механич. (тактильных), акустич., электрич. или оптич. раздражителей (сигналов) и их локации, т. е. определения положения по отношению к животному (см. Биолокация). Работа большинства механизмов локации обеспечивается парностью органов чувств (зрения, слуха, равновесия и др.), позволяющей сравнивать сигналы, сопоставляя силу, частотную характеристику и др. параметры сигналов, поступивших в правый и левый органы чувств, и т. о. определять направление их источника. Лит.: Протасов В. Р., Биоакустика рыб, М., 1965; Бионика. [Сб. ст.], М., 1965; Мазохин-Поршняков Г. А., Зрение насекомых, М., 1965; Глезер В. Д., Механизмы опознания зрительных образов, М.- Л., 1966; Райт Р. X., Наука о запахах, пер. с англ.. М., 1966; Мили Л. Дж. и М и л н М. Д ж., Чувства животных и человека, пер. с англ., М., 1966; С л о н и м А. Д., Инстинкт загадки врожденного поведения организмов, Л., 1967; Вопросы бионики. [Сб. ст.], М., 1967; Мартека В., Бионика, пер. с англ., М., 1967; Протасов В. Р., Зрение и ближняя ориентация рыб, М., 1968; Тинберген Н., Поведение животных, пер. с англ., М., 1969. Н. П. Наумов. БИОПОЛИМЕРЫ, высокомолекулярные природные соединения, являющиеся структурной основой всех живых организмов и играющие определяющую роль в процессах жизнедеятельности. К Б. относятся белки, нуклеиновые кислоты и полисахариды; известны также смешанные Б.- гликопротеиды, липопротеяды, гликолипиды и др. Биологические функции Б. Нуклеиновые кислоты выполняют в клетке генетич. функции. Последовательность мономерных звеньев (нуклеотидов) в дезоксирибонуклеиновой кислоте -ДНК (иногда в рибонуклеиновой кислоте - РНК) определяет (в форме генетического кода) последовательность мономерных звеньев (аминокислотных остатков) во всех синтезируемых белках и, т. о., строение организма и протекающие в нём оиохим. процессы. При делении каждой клетки обе дочерние клетки получают полный набор генов благодаря предшествующему самоудвоению (репликации) молекул ДНК. Генетич. информация с ДНК переносится на РНК, синтезируемую на ДНК как на матрице (транскрипция). Эта т. н. информационная РНК (и-РНК) служит матрицей при синтезе белка, происходящем на особых органоидах клетки - рибосомах (трансляция) при участии транспортной РНК (т-РНК). Биол. изменчивость, необходимая для эволюции, осуществляется на молекулярном уровне за счёт изменений в ДНК (см. Мутация). Белки выполняют в клетке ряд важнейших функций. Белки-ферменты осуществляют все химич. реакции обмена веществ в клетке, проводя их в необходимой последовательности и с нужной скоростью. Белки мышц, жгутиков микробов, клеточных ворсинок и др. выполняют сократит, функцию, превращая химич. энергию в механич. работу и обеспечивая подвижность организма в целом или его частей. Белки - осн. материал большинства клеточных структур (в т. ч. в специальных видах тканей) всех живых организмов, оболочек вирусов и фагов. Оболочки клеток являются липопротеид-ными мембранами, рибосомы построены из белка и РНК и т. д. Структурная функция белков тесно связана с регуляцией поступления различных веществ в субклеточные органеллы (активный транспорт ионов и др.)и с ферментативным катализом. Белки выполняют и регуляторные функции (репрессоры), "запрещая" или "разрешая" проявление того или иного гена. В высших организмах имеются белки - переносчики тех или иных веществ (напр., гемоглобин - переносчик молекулярного кислорода) и иммунные белки, защищающие организм от чужеродных веществ, проникающих в организм (см. Иммунитет). Полисахариды выполняют структурную, резервную и нек-рые др. функции. Белки и нуклеиновые к-ты образуются в живых организмах путём матричного ферментативного биосинтеза. Имеются теперь и биохим. системы внеклеточного синтеза Б. с помощью ферментов, выделенных из клеток. Разработаны методы химич. синтеза белков и нуклеиновых к-т. Первичная структура Б. Состав и последовательность мономерных звеньев Б. определяют их т. н. первичную структуру. Все нуклеиновые к-ты являются линейными гетерополимерами-сахарофосфатными цепочками, к звеньям к-рых присоединены боковые группы -азотистые основания: аденин и тимин (в РНК - урацил), гуанин и цитозин; в нек-рых случаях (гл. обр. в т-РНК) боковые группы могут быть представлены др. азотистыми основаниями. Белки -также гетерополимеры; молекулы их образованы одной или неск. полипептидными цепочками, соединёнными дисульфид-ными мостиками. В состав полипептидных цепей входит 20 видов различных мономерных звеньев - остатков аминокислот. Мол. масса ДНК варьирует от неск. млн. (у мелких вирусов и бактериофагов) до ста млн. и более (у более крупных фагов); бактериальные клетки содержат по одной молекуле ДНК с мол. массой в неск. млрд. ДНК высших организмов может иметь и большую мол. массу, ноизмерить её пока не удалось из-за разрывов в молекулах ДНК, возникающих при их выделении. Рибосомные РНК имеют мол. массу от 600 тыс. до 1,1 млн., информационная (и-РНК) - от сотен тысяч до неск. миллионов, транспортная (т-РНК) - ок. 25 тыс. Мол. масса белков варьирует от 10 тыс. (и менее) до миллионов; в последнем случае, однако, обычно возможно разделение белковой частицы на субъединицы, соединённые между собой слабыми, б. ч. гидрофобными, связями. К о н ф о р м а ц и я, т. е. та или иная пространственная форма молекул Б., определяется их первичной структурой. В зависимости от хим. строения и внеш. условий молекулы Б. могут находиться либо в одной или в неск. преимущественных конформациях (обычно встречающиеся в природных условиях нативные состояния Б.: напр., глобулярное строение белков, двойная спираль ДНК), либо принимать многие б. или м, равновероятные конформации. Белки делят по пространственной структуре на фибриллярные (нитевидные) и глобулярные; белки-ферменты, белки-переносчики, иммунные н нек-рые др. имеют, как правило, глобулярную структуру. Для ряда белков -гемоглобин, миоглобин, лизоцим, рибонуклеаза и др.- эта структура установлена во всех деталях (с определением при помощи рентгеноструктурного анализа расположения каждого атома). Она определяется последовательностью аминокислотных остатков и образуется и поддерживается относительно слабыми взаимодействиями между мономерными звеньями полипептидных цепей в водносолевом растворе (кулоновские и диполь-ные силы, водородные связи, гидрофобные взаимодействия), а также дисульфидными связями. Глобула белка формируется так, что большинство полярных гидрофильных аминокислотных остатков оказывается снаружи и контактирует с растворителем, а большинство неполярных (гидрофобных) остатков находится внутри и изолировано от взаимодействия с водой. Молекулы белка, обладающие избытком неполярных групп, когда часть из них оказывается на поверхности глобулы, образуют высшую, т. ц. четвертичную структуру, при к-рой неск. глобул агрегируют, взаимодействуя между собой в основном неполярными участками (рис. 1). Пространств, структура каждого белка-фермента уникальна и обеспечивает необходимое для его функционирования расположение в пространстве всех звеньев Б., в особенности т. н. активных центров. В то же время она не абсолютно жестка и допускает необходимые в процессе функционирования (при взаимодействии с субстратами, ингибиторами и др. веществами) конформационные сдвиги и изменения.
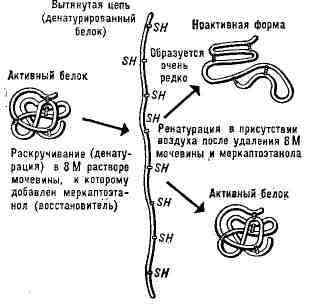
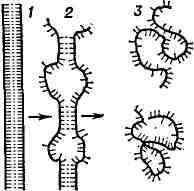
Пространств, структура нативной ДНК образована двумя комплементарными нитями и представляет собой двойную спираль Крика - Уотсона; в ней противоположные азотистые основания попарно связаны водородными связями - аденин с тимином и гуанин с цитозином. Устойчивость двойной спирали обеспечивается, наряду с водородными связями, также гидрофобным взаимодействием между плоскими кольцами азотистых оснований, расположенных стопкой (стопочное взаимодействие, или стакинг). Нити РНК спирализованы лишь частично. ДНК вирусов, бактериофагов, бактерий а также митохондриальная в ряде случаев представляет собой замкнутое кольцо; при этом наряду со спиралью Крика - Уотсона наблюдается ещё дополнительная т. н. сверхспирализация. Денатурация Б. Нарушение нативной пространств, структуры Б. при различных воздействиях (повышение темп-ры, изменение концентрации металлов, кислотности раствора и др.) наз. денатурацией и в ряде случаев обратимо (обратный процесс наз. ренатурацией; рис. 2). Молекулы Б.- кооперативные системы; поведение их зависит от взаимодействий составляющих частей. Коопера-тивность молекул Б. определяется тем, что повороты отдельных звеньев из-за внутримолекулярных взаимодействий зависят от конформации соседних звеньев. В основе денатурации Б. при изменении внешних условий обычно лежат кооперативные конформационные превращения (напр., переходы сс-спираль - b-структура, а-спираль - клубок, b-структура -клубок для полипептидов, переход глобула - клубок для глобулярных белков, переход спираль - клубок для нуклеиновых к-т). В отличие от фазовых переходов (кипение жидкости, плавление кристалла), являющихся предельным случаем кооперативных процессов и происходящих скачком, кооперативные переходы Б. совершаются в конечном, хотя и сравнительно узком, интервале изменений внешних условий. В этом интервале одномерные, линейные молекулы (нуклеиновые к-ты,полипептиды), претерпевающие переход спираль - клубок, разбиваются на чередующиеся спиральные и клубкообразные участки (рис. 3).
Рис. 2. Схема денатурации и ренатурации глобулярного белка (на примере фермента рибонуклеазы).
Рис. 3. Схема перехода спираль - клубок
для ДНК: 1 -нативное состояние (вместо двойной спирали для простоты изображена
"верёвочная лестница"); 2 - состояние ДНК в области перехода; 3 - денатурированное
состояние (однонитевые клубки).
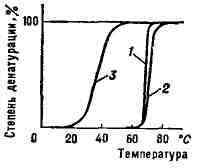
Рис. 4. Кривые перехода спираль - клубок (денатурации) нуклеиновых кислот из различных организмов: 1 - бактериальная ДНК; 2 - ДНК из зобной железы телёнка; 3 - РНК вируса табачной мозапки. Переход спираль - клубок в ДНК наблюдается при повышения темп-ры, добавлении в раствор к-ты или щёлочи, а также под влиянием др. денатурирующих агентов. Этот переход в гомополи-нуклеотидах происходит при нагревании в интервале десятых долей °С, в фаговых и бактериальных ДНК - в интервале 3-5°С (рис. 3), в ДНК высших организмов - в интервале 10-15°С. Чем выше гетерогенность ДНК, тем шире интервал перехода и меньше способность молекул ДНК к ренатурации. Переход спираль - клубок в различных видах РНК носит менее кооперативный характер (рис. 4) и происходит в более широком интервале темп-рных или др. денатурирующих воздействий. Б.- полимерные электролиты, их пространственная конформация и кооперативные переходы зависят как от степени ионизации молекулы, так и от концентрации ионов в среде, что влияет на электростатич. взаимодействия как между отдельными частями молекулы, так и между Б. и растворителем. Строение и биологи ч. функции Б. Строение Б.- результат длит, эволюции на молекулярном уровне, вследствие чего эти молекулы идеально приспособлены к выполнению своих биологич. задач. Между первичной структурой, конформацией Б. и конформационными переходами, с одной стороны, и их биол. функциями - с другой, существуют тесные связи, исследование к-рых - одна из гл. задач молекулярной биологии. Установление таких связей в ДНК позволило понять осн. механизмы репликации, транскрипции и трансляции, а также мутагенеза и нек-рых др. важнейших биологич. процессов. Линейная структура молекулы ДНК обеспечивает запись генетич. информации, её удвоение при матричном синтезе ДНК и получение (также путём матричного синтеза) мн. копий с одного и того же гена, т. е. молекул и-РНК. Сильные ковалентные связи между нуклеотидами обеспечивают сохранность генетич. информации при всех этих процессах. В то же время относительно слабые связи между нитями ДНК и возможность вращения вокруг простых химич. связей обеспечивают гибкость и лабильность пространств. структуры, необходимые для разделения нитей при репликации и транскрипции, а также подвижность молекулы и-РНК, служащей матрицей при биосинтезе белка (трансляция). Исследование пространств, структуры и конформационных изменений белков-ферментов на разных стадиях ферментативной реакции при взаимодействии с субстратами и коферментами даёт возможность установить механизмы биокатализа и понять природу огромного ускорения хим. реакций, осуществляемого ферментами. Методы исследования Б. При исследовании строения и конфор-мацнонных превращений Б. широко используются как очищенные природные Б., так и их синтетич. модели, к-рые проще по строению и легче поддаются исследованию. Так, при изучении белков моделями служат гомогенные или гетерогенные полипептиды (с заданным или случайным чередованием аминокислотных остатков). Моделями ДНК и РНК являются соответствующие синтетич. гомогенные или гетерогенные полинуклеотиды. К методам исследования Б. и их моделей относятся рентгеноструктурный анализ, электронная микроскопия, изучение спектров поглощения, оптич. активности, люминесценции, методы светорассеяния и динамич. двойного лучепреломления, седиментационный метод, вискозиметрия, физико-химич. методы разделения и очистки и ряд др. Все методы, разработанные для изучения синтетич. полимеров, применимы и к Б. При трактовке свойств Б. и их моделей, закономерностей их конформационных превращений используются также методы теоретич. физики (статистич. физики, термодинамики, квантовой механики и др.). Лит.: Б р е с л е р С. Е.. Введение в молекулярную биологию, М. - Л., 1966; В о л ь к е н ш т е й н М. В., Молекулы и жизнь, М., 1965; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967; Физические методы исследования белков и нуклеиновых кислот, М., 1967. Ю. С. Лазуркин. БИОПСИЯ (от био... к греч. opsis -вид, зрелище), иссечение кусочка болезненно изменённой ткани живого организма с последующим микроскопич. исследованием его для определения характера патологического процесса (воспаление, опухоль и т. д.). Б. позволяет не только уточнить клинич. диагноз, но и установить границы поражения. БИО - САВАРА ЗАКОН - закон, определяющий
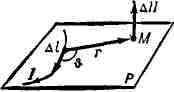
напряжённость магнитного поля, создаваемого электрич. током. Б.- С. з.
был открыт франц. учёными Ж. Б. Био (J. В. Biot) и Ф. Саваром (F. Savart)
в 1820 и сформулирован в общем виде П. Лапласом (P. Laplace). Согласно
этому закону, малый отрезок проводника дельта l (см. рис.), по к-рому
течёт ток силой I, создаёт в данной точке пространства М,
находящейся
на расстоянии r
от отрезка
магнитное поле напряжённостью
Здесь 0 - угол между направлением тока в отрезке дельта l и радиусом-вектором r, проведённым от отрезка к точке наблюдения М, a k - коэффициент пропорциональности, зависящий от выбора системы единиц. В системе СГС (Гаусса) k = 1/с, где с = 3*1010 см/сек - скорость света в вакууме, в системе СИ k = 1/4 Пи. Напряжённость магнитного поля ДЯ перпендикулярна плоскости Р, содержащей дельта l и r, и её направление определяется правилом буравчика: если вращать рукоятку буравчика (с правой нарезкой) от дельта l к r, то поступательное движение буравчика укажет направление дельта Н. Полная напряжённость магнитного поля Н, создаваемого проводником с током в точке М, равна векторной сумме величин дельта Н, обусловленных всеми элементами дельта l проводника. В частности, напряжённость Н магнитного поля на расстоянии d от длинного (много больше d) прямого провода, по к-рому течёт ток силой I, равна; H = k2I/d: в центре кругового контура (радиуса R), по которому течёт ток силой I, H = k*2ПиI/R, a на его оси в точке, отстоящей от плоскости контура на расстоянии d>>R, H = k*2ПиR2I/d3; на оси соленоида из я витков Н = k*4ПипI. Б.- С. з. можно рассматривать также как закон, определяющий магнитную индукцию ДВ. В системе СГС для этого нужно выражение для ДН умножить на магнитную проницаемость среды ц, а в системе СИ, кроме того,- на магнитную проницаемость вакуума n0 = 4Пи*10-7гн/м. Г. Я. Мякишев. БИОСИНТЕЗ (от био... и синтез), образование органич. веществ из более простых соединений, протекающее в живых организмах или .вне их под действием биокатализаторов - ферментов. Б.- часть процесса обмена веществ растений, животных и микроорганизмов. Непосредственным источником энергии для Б. служат богатые энергией соединения (см. Биоэнергетика), а в конечном счёте (для всех организмов, кроме бактерий, осуществляющих хемосинтез) -энергия солнечного излучения, аккумулированная зелёными растениями (см. Ассимиляция, Фотосинтез). Каждый одноклеточный организм, как и каждая клетка многоклеточного организма, синтезирует составляющие её вещества. Характер Б., осуществляемого в клетке, определяется наследств, информацией, "закодированной" в её генетич. аппарате (см. Белки, Биосинтез; Генетический код). Б., производимый вне организмов, широко применяется как способ (иногда единственно возможный) пром. получения биологически важных веществ - витаминов, нек-рых гормонов, антибиотиков, аминокислот, а также белков и др. соединений. См. Микробиологическая промышленность.. С.Е.Северин. БИОСИСТЕМАТИКА, раздел ботаники, изучающий таксономическую и популяционную структуру вида, его морфологогеографич., экологич. и генетич. дифференциацию, происхождение и эволюцию. Б. оперирует не только собственно таксономич. категориями, как вид и подвид, но и генэкологическими и популяционно-генетическими - экотып, биотип, популяция идем (элементарная локальная популяция) или г а м о д е м (у амфимиктич. растений). Б. возникла как наука, сочетающая различные подходы к структуре и эволюции вида, т. е. задачи её выходят за рамки собственно систематики. История Б. начинается с работ швед, эколога Г. Турессона (1922, 1923) и амер. эколога Д. Клаусена (1921-22), изучавших экологич. и генетич. дифференциацию вида. Новое направление, назв. Турессоном (1923) генэкологией, сформировалось позднее в науку, к-рую М. Кэмп и Н. Гилли назвали "Б." (1943). Генэкология осталась одним из разделов Б., изучающим внутривидовую изменчивость растений. Б. изучает, кроме того, и микроэволюцию. В СССР работы в этом направлении начали ещё в 20-х гг. М. А. Розанова, Е. Н. Синская и др. Под руководством Н. И. Вавилова во Всесоюзном институте растениеводства велось изучение экологогеографич. и генетич. дифференциации мн. видов культурных растений. Эти исследования имели большое значение для дальнейшего развития Б., хотя они и относились скорее к "дифференциальной систематике", как её понимал Н. И. Вавилов. Лит.: Вавилов Н. И., Линнеевскпй вид как система, "Тр. по прикладной ботанике, генетике и селекции" , 1931, т. 26, т. 3,с. 109-34; Розанова М. А., Экспериментальные основы систематики растений, М.-Л., 1946; Списка я Е. Н., Динамика вида, М.-Л., 1948; Завадский К. М., Вид и видообразование, Л., 1968; Тахтаджян А. Л., Биосистематика: прошлое, настоящее и будущее, "Ботанический журнал", 1970, т. 55, в. 3; Н е s 1 о р-Н arrison J. W., New concepts in flowering-plant taxonomy, L., 1953; его же. Forty years of Geneco-logy, в сб.: Advances in ecological research, y. 2, L.-N. Y., 1965; Da vis P. H. and Heywood V. H., Principles of angiosperm taxonomy, Edinburg-L., 1963: Reproductive biology and taxonomy of vascular plants, ed. J. G. Hawkes, Oxf., 1966; Modern methods in plant taxonomy, ed. V. H. Heywood, L., 1968; В r i g g s D. and Walters S. M., Plant variation and evolution, L., 1969. А. Л. Тахтаджян. БИОСТРАТИГРАФИЯ (от био... и стратиграфия), отрасль стратиграфии, изучающая распределение ископаемых остатков организмов в осадочных отложениях с целью установления относит, возраста и соотношения одновозрастных слоев на различных территориях. Задача Б.- разработка шкал относит, возраста слоев (разной детальности и масштаба, в частности зональных). Последовательность биостратиграфич. зон отражает смену в геологич. разрезе ископаемых остатков группы вымерших организмов разного систематич. ранга или их комплексов. Особенное значение для выделения зон, и в первую очередь биозон, имеют группы вымерших организмов с относительно кратким сроком существования, но достигавшие широкого распространения, значит, изобилия и разнообразия (напр., нуммулиты, граптолиты, динозавры). Нередко зоны обосновываются стадиями эволюции нек-рых быстро изменявшихся во времени групп вымерших организмов (напр., кораллов - ругоз). Для целей Б. важно изучение остатков древних микроскопич. организмов (микропалеонтология), количество к-рых может быть велико даже в небольших образцах (напр., из глубоких скважин). Остатки планктонных организмов (фора-минифер, водорослей и др.), разносившихся течениями на большие расстояния, допускают выделение зон большой территориальной протяжённости. Ископаемые остатки спор и пыльцы растений, далеко разносившихся ветрами, важны для корреляции одновозрастных осадков мор. и континент, происхождения. Б. широко использует методы палеоэкологии для реконструкции условий существования древних организмов, с тем чтобы отличать одновозрастные комплексы организмов, живших в разных условиях, от разновозрастных, живших в сходных условиях. Лит.: М е н н е р В. В., Биостратиграфические основы сопоставления морских, лагунных и континентальных свит, "Тр. геологического ин-та АН СССР", 1962, в. 65; Стратиграфическая классификация, терминология и номенклатура, Л., 1963; Степанов Д. Л.. Принципы и методы биостратиграфических исследований, Л., 1958. Р. Л. Мерклин. БИОСФЕРА (от био... и сфера), оболочка Земли, состав, структура и энергетика к-рой в существ, чертах обусловлены прошлой или совр. деятельностью живых организмов. Б. охватывает часть атмосферы, гидросферу и верхнюю часть литосферы, к-рые взаимосвязаны сложными биогеохимич. циклами миграции веществ и энергии (по В. И. Вернадскому,-биогенная миграция атомов); начальный момент этих циклов заключён в трансформации солнечной энергии растениями и синтезе биогенных веществ на Земле (см. Фотосинтез, Хемосинтез). Термин "Б." ввёл в 1875 австр. геолог Э. Зюсс. Общее учение о Б. создано в 20-30-х гг. 20 в. В. И. Вернадским, развившим идеи В. В. Докучаева о комплексном естествен-но-историч. анализе взаимодействующих в природе разнокачеств. объектов и явлений (факторов почвообразования) и выявлении самостоятельных природных объектов гетерогенной структуры и состава (почвы, природные зоны). В основе учения Вернадского лежат представления: I) о планетарной геохимич. роли живого вещества (совокупность всех живых организмов, существовавших или существующих в определённый отрезок времени, рассматриваемых как мощный геологич. фактор; в отличие от живых существ, изучаемых в биологии на всех уровнях их организации, начиная от молекулярного, живое вещество, в понимании Вернадского, как биогеохимич. фактор, количественно выражается в элементарном химич. составе, массе и энергии) и 2) об организованности Б., являющейся продуктом сложного превращения вещественно-энергетич. и информац. потоков живым веществом за время геологич. истории Земли. Б. включает не только область жизни (биогеосферу, фитогеосферу, геомериду, витасферу), но и др. структуры Земли, генетически связанные с живым веществом. По Вернадскому, вещество Б. состоит из семи разнообразных, но геологически взаимосвязанных частей: живое вещество; биогенное вещество; косное вещество; биокосное вещество; радиоактивное вещество; рассеянные атомы; вещество космич. происхождения. В пределах Б. везде встречается либо живое вещество, либо следы его биогеохимич. деятельности. Газы атмосферы (кислород, азот, углекислота), природные воды, равно как и каустобиолиты (нефти, угли), известняки, глины и их метаморфич. производные (сланцы, мраморы, граниты и др.) в своей основе созданы живым веществом планеты. Слои земной коры, лишённые в наст, время живого вещества, но переработанные им в геология, прошлом, Вернадский относил к области "былых биосфер". Б. мозаична по структуре и составу, отражая геохимич. и геофизич. неоднородность лика Земли (океаны, озёра, горы, ущелья, равнины и т. д.) и неравномерность в распределении живого вещества по планете как в прошлые эпохи, так и в наше время. Максимальное содержание живого вещества гидросферы приурочено к мелководьям, минимальное - к глубинным акваториям (абиссаль); на суше эта неравномерность проявляется в мозаике биогеоценотич. покрова (леса, болота, степи, пустыни и др.) с минимумом плотности живого вещества в высокогорьях, пустынях и полярных областях (см. Биомасса). Элементарная структура активной части совр. Б.- биогеоценоз. Живое вещество выполняет след, биогеохимич. функции: газовые (миграция газов и их превращения); концентрационные (аккумуляция живыми организмами химич. элементов из внешней среды); окислительно-восстановительные (химич. превращения веществ, содержащих атомы с переменной валентностью,- соединений железа, марганца, микроэлементов и т. д.); биохимические и биогеохимические функции, связанные с деятельностью человека (техногенез, форма созидания и превращения вещества в Б., стимулирующая переход Б. в новое состояние - ноосферу). Совокупность этих функций определяет все химич. превращения в Б. Эволюция Б. диалектически связана с эволюцией форм живого вещества (организмы и их сообщества), усложнением его биохимич. функций, совершающихся на фоне геологич. истории Земли. В учении о Б. выделяют след. осн. аспекты: энергетический, освещающий связь биосферно-планетарных явлений с космич. излучениями (в осн. солнечными) и радиоактивными процессами в земных недрах; биогеохимический, отражающий роль живого вещества в распределении и поведении атомов (точнее их изотопов) в Б. и её структурах (см. Биогеохимия); информационный, изучающий принципы организации и управления, осуществляемые в живой природе в связи с исследованием влияния живого вещества на структуру и состав Б.; пространственно-временной, освещающий формирование и эволюцию различных структур Б. в геологич. времени в связи с особенностями пространственно-временной организованности живого вещества в Б. (проблемы симметрии и др.); н о о с ф е р н ы и, изучающий глобальные эффекты воздействия человечества на структуру и химию Б.: разработка полезных ископаемых, получение новых, отсутствовавших до того в Б. веществ (напр., чистые алюминий, железо и др. металлы), преобразование биогеоценотических структур Б. (сведение лесов, осушение болот, распашка целинных земель, создание водохранилищ, загрязнение вод, почв и атмосферы продуктами хоз. деятельности, внесение удобрений, эрозия почв, лесонасаждение, строительство городов, плотин, промысловое х-во и т. д.). Выход человека в космос, за пределы Б., будет стимулировать разработку новых сторон учения о Б. Существ, момент учения о Б.- представления о взаимосвязях (прямых и обратных связях) и сопряжённой эволюции всех структур Б. Это представление положено в основу разработки мн. нац. и междунар. организациями, науч. центрами и лабораториями проблемы "биосфера и человечество". Решению этой проблемы служат мероприятия, в к-рых участвуют мн. страны, напр. Международное гидрологич. десятилетие, Международная биологич. программа (см. Биологическая программа международная) и т. д. Повышенный интерес к изучению Б. вызван тем, что локальное воздействие человека на Б., характерное для всей предшествовавшей истории, сменилось в 20 в. глобальным его влиянием на состав, структуру и ресурсы Б. На планете нет участка суши или моря, где бы не были обнаружены следы деятельности человека. Один из ярких примеров - глобальные выпадения радиоактивных осадков -продуктов ядерных взрывов. В атмосфере, океане и на суше повсеместно присутствуют (пусть в самых незначит. кол-вах) продукты сгорания нефти, угля, газов, отходы химич. и др. индустрии, ядохимикаты и удобрения, сносимые с полей в процессе водной и ветровой эрозии. Интенсивное и нерациональное использование ресурсов Б.- водных, газовых, биологич. и др., усугубляемое гонкой вооружений, испытаниями ядерного оружия и т. д., развеяло миф о бесконечности и неисчерпаемости этих ресурсов. Многочисленные примеры разрушит, деятельности человека и, к сожалению, редкие примеры его созидательной деятельности (в т. ч. в плане охраны природы) свидетельствуют об актуальности разумного ведения земных дел разумным человечеством, что возможно только при переходе от стихийного капиталистич. произ-ва к плановому х-ву социалистич. и коммунистич. общества. Естественно-науч. основой рационального подхода к проблеме "биосфера и человечество" -одной из грандиознейших проблем нашего времени - служат учение о Б. и биогеоценология - дисциплины, изучающие общие принципы и механизмы функционирования и эволюции сообществ живых организмов в определённых пространственных и временных условиях. Совр. структура Б.- продукт длительной эволюции мн. систем разной сложности, последовательно стремящихся к состоянию динамич. равновесия. Практич. значение учения о Б. огромно. Особенно заинтересованы в развитии этого учения здравоохранение, сельское ц промысловое х-во и др. отрасли человеч. практики, чаще других сталкивающиеся с "ответными ударами" со стороны Б., вызванными неразумным или неосторожным преобразованием природы человеком. Лит.: Вернадский В. И., Избр. соч., т. 5, М., 1960; его же, Химическое строение биосферы Земли п её окружения, М., 1965; К о в д а В. А., Современное учение о биосфере, "Журнал общей биологии", 1969, т. 30, № 1; П е р е л ь м а н А. И., Геохимия ландшафта, М., 1961; Тимофеев-Ресовский Н. В. и Т ю р ю к а н о в А. Н., Об элементарных биохорологическнх подразделениях биосферы, "Бюллетень Московского общества испытателей природы", 1966, т. 71(1); X н л ь м и Г. Ф., Основы физики биосферы, Л., 1966; Д ю-в и н ь о П. и Ганг М., Биосфера и место в ней человека,.пер. с франц.,М., 1968. В. А. Ковда, А. Н. Тюрюканов. БИОТА (от греч. biote - жизнь), исторически сложившаяся совокупность растений и животных, объединённых общей областью распространения. В отличие от биоценоза, виды, входящие в состав Б., могут и не иметь экологич. связей (напр., кенгуру и двоякодышащая рыба цератодус, входящие в состав австрал. фауны). Однако во мн. случаях одна и та же совокупность организмов может рассматриваться и как Б. (с позиций биогеографии) и как биоценоз (с позиций экологии). Лит.: Б о б р н н с к н и Н. А., География животных. М.. 1951. БИОТА (Thuja), род однодомных древесных растений сем. кипарисовых. Представлен 1 видом - Б. восточной (Thuja orientalis, Biota orientalis). Биота восточная: а - ветвь с женскими шишками; б - веточка. Дерево выс. 8-10 м, но чаще кустарник. Хвоя на взрослых ветвях чешуевидная, накрест-супротивно расположенная. Крона яйцевидная, состоит из мн. плоских побегов ("пластин"), расположенных в вертикальной плоскости. Направленные вверх шишки незрелые -голубовато-зелёные, впоследствии - сухие, б. ч. красновато-коричневыс. Семена созревают на второй год. Родина Б.- Китай и Корея. В юж. районах СССР разводится как декоративное растение. Б. засухоустойчива, хорошо выносит стрижку. Лит.: Деревья и кустарники СССР, т. 1, М. - Л., 1949. А. П. Шиманюк. БИОТЕЛЕМЕТРИЯ (от био..., теле... и ...метрия), способ дистанционного исследования биологич. явлений и измерения биологич. показателей. При Б. на изучаемом объекте (животном или человеке) укрепляют соответствующие датчики, сигналы к-рых, характеризующие те или иные биол. или физиологич. процессы (движение, пульс, дыхание и др.), передают по каналам связи (радио- или телефонная связь) и регистрируют на пункте приёма информации. Исследуемый процесс, если он неэлектрич. природы, предварительно преобразуют в к.-л. электрич. сигналы. Применение телеметрии даёт возможность проводить исследование на очень больших расстояниях (напр., при космич. полётах) или во время движения изучаемого объекта (напр., во время спортивных соревнований или трудовой деятельности). При Б. возможна передача сигналов и о процессах, происходящих во внутр. органах, для чего один или неск. сверхминиатюрных радиопередатчиков (т. н. радиокапсул) вводят в полости тела (напр., в желудок или кишки, см. Эндорадиозондированир) пли вживляют в ткани организма. Посредством Б. можно изучать поведенческие реакции животных в обычной для них среде обитания и в таких условиях, в к-рых прежде было невозможно исследование физиологии, процессов, напр, при полёте птиц. Б. приобрела важное значение в кос-мич. биологии и космич. медицине, в физиологии труда и спорта, а также в экологии и физиологии животных. См. также Телеуправление, Телесигнализация. Лит.: Биотелеметрия. [Сб. ст.], пер. с англ., М., 1965; Розенблат В. В., Радиотелеметрические исследования в спортивной медицине, М., 1967; Проблемы радиотелеметрии в физиологии и медицине. Материалы III Всесоюзного симпозиума, Свердловск, 1968; Caceres С. A., Cooper I. К., Biomedical telemetry, N. Y. -L., 1965. Е.Б.Бабский. БИОТЕРМИЧЕСКАЯ ЯМА, Беккари яма, пирятинская яма, чешская яма, сооружение для уничтожения трупов животных. Строится по типовому проекту из влаго-и термоустойчивого материала, имеет герметич. крышку и отверстия для притока воздуха. Через 20 сут после загрузки трупами темп-pa в камере поднимается до 65° С. Процесс разложения трупов заканчивается за 35-40 сут с образованием однородного не имеющего запаха компоста, пригодного для удобрения.Б. я. имеют значит, преимущество перед скотомогильниками, т. к. обеспечивают быструю гибель многих микробов. БИОТЕХНИЧЕСКАЯ СИСТЕМА, совокупность взаимосвязанных и взаимозависимых биологич. и технич. систем или объектов. Напр, на борту космич. корабля Б. с. состоит из подобранного, в зависимости от назначения и продолжительности полёта, биокомплекса и технич. средств, обеспечивающих оптимальные условия его функционирования. В состав технич. средств входят подсистемы создания и распределения света, энергообеспечения, терморегулирования, а также космич. оранжерея, кухня, блоки регенерации воздуха и воды, минерализации отходов и т. д. Примерами Б. с. могут служить также электростимулятор сердца, манипулятор для работы в условиях, при к-рых соприкосновение человека с объектом управления нежелательно, и т. д. (см. Система "человек и машина"). БИОТЕХНИЯ (от био... и греч. recline -умение, мастерство), комплекс мероприятий, направленных на увеличение запасов полезных животных и улучшение их продуктивных свойств. Термин "Б." появился в 30-х гг. 20 в. в СССР, где широко развернулись работы по охране и разведению в природных условиях промысловых животных. Организация действенной охраны (путём создания заповедников, заказников и гос. заповедно-охотничьих хозяйств, установления сроков добычи животных, запрета добычи на определённый срок и др. мер) и проведение биотехнич. мероприятий позволили восстановить и увеличить численность многих ценных видов зверей, птиц и рыб, почти полностью истреблённых хищнич. промыслом в дореволюц. России. Биотехнич. мероприятия имеют целью: 1) увеличение запасов промысловых животных, улучшение кормовой базы (посадка и посев кормовых растений в охотничьих угодьях и водоёмах, подкормка животных); улучшение защитных и гнездовых условий (посадка деревьев и кустарников, посевы высокостебельчатых трав для укрытия животных; устройство защитных участков - ремиз, обсаженных колючим кустарником); создание участков "покоя" в глухих малопосещаемых местах; устройство нор и др. убежищ для разных видов животных; истребление вредных хищников; борьба с заболеваниями животных; реакклиматизация зверей, птиц и рыб в районах прежнего обитания; внедрение в фауну страны ценных диких животных, завезённых из др. стран; 2) улучшение продуктивных свойств промысловых животных (повышение меховых достоинств пушных зверей и мясных качеств копытных животных и пернатой дичи, увеличение размеров тела добываемых рыб и др.) включает селекцию (отбор животных в соответствии с поставленными задачами и подбор родительских пар с целью усиления в потомстве полезных признаков, метизацию и гибридизацию, использование мутационных форм с ценными продуктивными свойствами), а также создание условий, способствующих развитию продуктивных свойств животных. Большой вклад в Б. внесли сов. учёные: Б. М. Житков, П. А. Мантейфель, С. И. Огнев, Н. П. Лавров, С. П. Наумов, Б. А. Кузнецов, И. Н. Арнольд, А. Н. Елеонский, В. П. Врасский, В. А. Мовчан, Г. В. Никольский и др. В СССР разработкой науч. проблем Б. и внедрением их в практику охотничьего хозяйства занимается Всесоюзный н.-и. ин-т охотничьего хозяйства и звероводства и др. научные учреждения. Большой опыт в разработке и осуществлении различных биотехнич. мероприятий накоплен в охотничьем и рыбном хозяйствах многих зарубежных стран (США, Англии, Франции, Канады, Финляндии, Югославии, Венгрии, Чехословакии, ГДР и др.). См. Охотничье хозяйство, Охота. Лит.: Колосов А. М., Биотехния, М., 1965; Дементьев В. И., Биотехнические мероприятия в охотничьем хозяйстве, Л., 1966; Кузнецов Б. А., Биотехнические мероприятия в охотничьем хозяйстве М., 1967; Колосов А. М., Лавров Н. П., Обогащение промысловой фауны СССР, М., 1968. Б. А. Кузнецов. БИОТИН (витамин Н), водорастворимый
витамин, содержащийся в дрожжах и др. микроорганизмах, а также в печени,
яичном желтке, почках, молоке, цветной капусте и др. растит, продуктах.
Суточная потребность человека Б. (0,25 мг) обычно обеспечивается
микрофлорой кишечного тракта, поэтому вводить его с пищей не нужно. Б.
получен синтетически.
У лабораторных животных можно вызвать авитаминоз Н скармливанием больших количеств сырого яичного белка, содержащего антагонист Б.- авидин, разрушающийся при нагревании до 100°С. При авитаминозе Н наблюдаются поражения кожи типа себореи. Б.- кофермент, участвует в процессах переноса углекислого газа и фиксации его при биосинтезе; др. функции его не выяснены. БИОТИП, группа организмов, входящих в состав местной популяции, имеющих одинаковый генотип и сходных практически по всем признакам. Дат. биолог В. Иогансен считал гомозиготный Б. у самоопыляющихся растений самой элементарной единицей структуры популяции (1909). Сов. ботаник В. Н. Сукачёв (1927, 1935) называл Б. клоны растений, полученные многократным черенкованием корней растения или делением одного куста. В популяциях перекрёстноопыляемых видов Б. можно выделить как последовательным отбором с изоляцией потомства, так и многократным размножением в близких степенях родства (инбридинг). В 20-30-е гг. 20 в. мн. биологи (швед, ботаник Г. Турессон, сов. биологи Н. И. Вавилов, М. А. Розанова и др.) считали Б. мельчайшей таксономия, единицей. Вид, по их мнению, складывается из совокупности Б. в результате перекомбинирования и отбора. Биологическими типами, или жизненной формой, наз. также большие группы организмов, часто состоящие из многих видов и характеризующиеся сходными приспособлениями к использованию определённых условий обитания. К биологическим типам в этом значении термина относят, напр., группу подземных роющих грызунов, группу эфемерных растений пустынь и т. д. Лит.: Б е р м а н 3. И., Завалский К. М., 3 е л и к м а н А. Л. и др., Современные проблемы эволюционной теории, Л., 1967; Завадский К. М., Вид и видообразование, Л., 1968. К.М.Завадский. БИОТИТ [по имени франц. учёного Ж. Б. Био (J. В. Biot), 1774-1862], минерал из группы слюд. По структуре относится к слоистым алюмосиликатам. Химич. формула K(Mg,Fe)3AlSi3O10(OH,F)2. Цвет в тонких листочках от черновато-бурого до буро-зелёного. Б. широко распространён как породообразующий минерал в изверженных и метаморфических породах. Наиболее крупные кристаллы Б., достигающие 1-1,5 м, встречаются в пегматитовых жилах. Б. применяют в малоответственных электроизоляционных изделиях, порошок его также идёт на изготовление бронзовой краски. См. Слюды. БИОТИЧЕСКИЕ ФАКТОРЫ среды, совокупность влияний, оказываемых на организмы жизнедеятельностью др. организмов. Эти влияния носят самый разнообразный характер. Живые существа могут служить источником пищи для др. организмов, являться средой обитания (напр., организм-хозяин, в к-ром поселяются паразиты), способствовать их размножению (напр., деятельность животных-опылителей), оказывать химич. (токсины бактерий), механич. и др. воздействия. В отличие от абиотических факторов среды, действие Б. ф. проявляется в форме взаимовлияния живых организмов разных видов друг на друга. Так, растения выделяют кислород, необходимый для дыхания животных, а животные обеспечивают поступление в атмосферу углекислого газа, к-рый используется растениями в процессе фотосинтеза; деятельность хищников оказывает влияние на динамику численности их жертв, что, в свою очередь, сказывается на изменениях численности хищников. Действие Б. ф. может быть не только непосредственным, но и косвенным, выражаясь в изменении условий окружающей неживой природы (напр., изменение состава почвы бактериями или изменение микроклимата под пологом леса). Лит.: Наумов Н. П., Экология животных, М., 1955; Макфедьен Э., Экология животных. Цели н методы, [пер. с англ.], М., 1965. И. А. Шилов. БИОТИЧЕСКИЙ ПОТЕНЦИАЛ в экологии, способность вида противостоять неблагоприятным воздействиям внешней среды. Термин введён амер. экологом Р. Чепменом (1925) в связи с проблемой динамики численности животных. По Чепмену, Б. п.- количественное выражение способности организмов противостоять сопротивлению внешней среды. Согласно его теории, потенциальная плодовитость животных не реализуется, поскольку она подавляется односторонним воздействием внеш. среды, с к-рой организмы находятся в антагонистич. отношениях. По совр. воззрениям, такая точка зрения выглядит упрощённой. Изменения плодовитости и выживания животных происходят как под влиянием абиотич. факторов, так и в результате межвидовых и внутривидовых взаимоотношений. Большую роль в этих процессах играют внутрипопуляционные механизмы, обеспечивающие активную реакцию популяции на внешние воздействия. Лит.: Наумов Н. П., Экология животных, М., 1955; В и л л и К., Биология, пер. с англ., М., 1968, с. 700. И.А.Шилов. БИОТОП (био... и греч. topos - место), участок земной поверхности (суши или водоёма) с однотипными абиотич. условиями среды (рельеф, почвы, климат и т. п.), занимаемый тем или иным биоценозом. Характерный для данного Б. комплекс условий определяет как видовой состав организмов, так и особенности их существования и, в свою очередь, подвергается изменениям под воздействием биоценоза. Т. о., Б.- неорганический компонент биогеоценоза. Сходные Б. объединяют в биохоры, совокупности к-рых составляют биоциклы. БИОТОПЛИВО, биологическое топливо, различные органич. материалы, выделяющие в процессе разложения тепло, которое используется для обогрева теплиц, парников и утеплённого грунта. В качестве Б. применяют навоз (конский, коровий, овечий, свиной), бытовой мусор, корьё (кора, снятая с дерева), древесные опилки, льняную костру, отходы текстильной пром-сти, сухой древесный лист, неразложивш-ийся торф. При ср. плотности укладки объёмная масса составляет (в т/м3): навоза конского и овечьего 0,35-0,45, коровьего-0,40-0,50, бытового мусора - 0,70 -0,75, корья - 0,40,-0,45. Б. поздней осенью складывают на хранение (раздельно по видам) в штабели, сильно утрамбовывая и утепляя соломистым навозом. Во время хранения в Б. поддерживают температуру от 0 до 10°С. За 2-3 нед до использования Б. перебивают (разрыхляют). Для ускорения разогревания сырое и плотное Б. смешивают с сухим и рыхлым. Если в течение 1 нед Б. не разогревается, прибегают к его искусств, разогреву (укладка очагами горячего Б. или негашёной извести, укладка горячих камней и т. д.). На 1 м2 площади теплиц и утеплённого грунта требуется Б. 0,25 -0,4 м3, на 1 рамоместо парников 0,6 -1,5 м3. В. А. Брызгалов. БИОУПРАВЛЕНИЕ, система управления приборами, механизмами и устройствами, в к-рой в качестве управляющих сигналов используются различные проявления жизнедеятельности организма, за исключением большинства произвольных движений. Для Б. могут служить: биоэлектрические потенциалы, генерируемые различными возбудимыми тканями, механич. и акустич. явления, сопровождающие функционирование сердечно-сосудистой системы и дыхания, колебания темп-ры тела и др. Наиболее широко распространены системы биоэлектрического управления. В этих системах биопотенциалы, генерируемые скелетными мышцами, сердцем, головным мозгом, нервами, подвергаются усилению, переработке и затем выполняют роль командных, управляющих сигналов. Использование биопотенциалов головного мозга позволило создать приборы для автоматич. сигнализации начальной стадии кислородного голодания, для автоматич. управления подачей наркотич. вещества и поддержания заданной стадии наркоза, прибор для автоматич. управления электроэнце-фалографом (см. Электроэнцефалография) в связи с выделением характерных изменений состояния мозга. Больше всего приборов, управляемых биопотенциалами сердца. При этом в качестве сигнала могут служить, например, характерные изменения электрокардиограммы при заболеваниях. Первая группа приборов, управляемых биопотенциалами сердца, это приборы диагностические, обеспечивающие включение сигнализирующей и регистрирующей аппаратуры при нарушениях сердечного ритма, кислородном голодании сердечной мышцы и др. Вторая группа - приборы лечебного назначения, служащие для автоматяч. включения электростимулятора, задающего нужный ритм сердечных сокращений (при нарушениях естеств. ритма, резком замедлении сердечных сокращений или остановке сердца), для осуществления синхронного с сердечными сокращениями массажа периферич. сосудов, для временной разгрузки сердца с помощью вспомогательного искусственного сердца. Важную группу устройств с биоэлектрич. управлением составляют активные протезы, для управления к-рыми используются биопотенциалы частично ампутированных, парализованных или полностью сохранённых мышц. Выполняя привычные движения, человек управляет электромеханич. или пневматич. приводом, к-рый осуществляет движения в суставах парализованной конечности или шарнирах протеза. В 60-х гг. 20 в. не только в СССР, но и в Англии, Канаде (по сов. лицензиям) налажен пром. выпуск биоуправляемых протезов. Биоэлектрическое управление применяют также в технике, напр., в биоманипуляторах, управляемых на расстоянии при работе в подводных или вредных условиях. Лит.: Кобринский А. Е. [и др.], Биоэлектрическая система управления, "Докл. АН СССР", 1957, т. 117, № 1; Гурфинкель В. С., Биоэлектрическое управление в медицине, "Вести. АМН СССР", 1964, № 2. В. С. Гурфинкель. БИОФАБРИКА, гос. хозрасчётное предприятие, изготовляющее биологич. препараты для диагностики, профилактики и лечения болезней животных. В СССР Б. начали создавать с 1930 на базе вет. бактериологич. лабораторий и станций по производству вакцин и сывороток. Вначале Б. были небольшой производительности с узким ассортиментом выпуска биологических препаратов. В 1970 большинство Б. представляет собой предприятия широкого профиля, изготовляющие 8-10 видов биологических препаратов. БИОФИЗИКА, биологическая физика, наука, изучающая физич. и физико-химич. процессы, протекающие в живых организмах, а также ультраструктуру биологич. систем на всех уровнях организации живой материи - от субмолекулярного и молекулярного до клетки и целого организма. Развитие Б. тесно связано с интенсивным взаимопроникновением идей, теоретич. подходов и методов современной биологии, физики, химии и математики. Развитие биологии показало, что для понимания и изучения элементарных биологич. явлений необходимо применение понятий и методов точных наук. Такой подход оправдан тем, что все биологич. объекты представляют в конечном итоге совокупность атомов и молекул и подчиняются физич. и химич. закономерностям. Но так как биологич. системы - это самоорганизующиеся системы, сложившиеся в процессе эволюции, им присущи многие свойства, не имеющие места в неживой природе. Сложность биологич. систем обеспечивает протекание процессов, маловероятных для условий, обычно рассматриваемых в физике. Б. в основном рассматривает целостные системы, не разлагая их, по возможности, на химич. компоненты. В связи с этим возникает необходимость перерабатывать известные физико-химич. методы, создавая высокоспециализированные биофизич. методы и приёмы. Совр. Б., согласно классификации, принятой Международным союзом теоретич. и прикладной биофизики (1961), включает следующие осн. разделы: молекулярная Б., в задачу к-рой входит исследование физич. и физико-химич. свойств макромолекул и молекулярных комплексов, составляющих живые организмы, а также характера взаимодействия и энергетики протекающих в них процессов; Б. клетки, изучающая физико-химич. основы функции клетки, связь молекулярной структуры мембран и клеточных органелл с их функцией, механич. и электрич. свойства, энергетику и термодинамику клеточных процессов; Б. процессов управления и регуляции, к-рая занимается исследованием и моделированием внутр. связей системы управления в организмах, их физич. природой, исследованием физич. закономерностей живого на уровне целого организма. Однако исторически сложившийся круг проблем, к-рыми занимается Б., шире. К Б. относится: изучение влияния физич. факторов на организм (см. Вибрация, Ускорение, Невесомость); исследование биологического действия ионизирующих излучений, к-рое в связи с важностью и актуальностью этого вопроса стало предметом радиобиологии, специальной науки, выделившейся из Б. Физич. анализ деятельности органов чувств, в первую очередь оптики глаза, анализ работы органов движения, дыхания, кровообращения как физич. систем, вопросы прочности и эластичности тканей (см. Биомеханика) - существенные, исторически сложившиеся разделы Б. Важное значение имеет и разработка физич. методов исследования биологич. систем - от макромолекул до целого организма, без к-рых невозможно совр. биологич. исследование. Отдельные исследования биофизич. характера можно проследить с 17 в. В этот период были сделаны попытки применить понятия, созданные в физике и химии, для анализа биологич. явлений. Франц. учёный Р. Декарт рассматривал человеческое тело как сложную машину. Он опубликовал ряд работ по исследованию органов чувств - биоакустике и оптике. Последователь Декарта - итал. учёный Дж. А. Борелли пытался объяснять движение живых существ чисто физич. закономерностями. Л. Эйлер, проф. Петербургского ун-та, впервые математически описал движение крови по сосудам. М. В. Ломоносов выдвинул в 1756 одну из первых гипотез цветного зрения. Могучим толчком к физико-химич. исследованиям явлений жизни послужили опыты итал. учёного Л. Гальвани, к-рый доказал наличие "животного электричества". Во 2-й пол. 19 в. нем. учёные Г. Гельмгольц и В. Вундт сформулировали осн. закономерности физиологической акустики и физиологической оптики. Нем. врач Ю. Р. Майер, наблюдая насыщение кислородом гемоглобина в крови человека в тропич. и умеренном климате, сформулировал закон сохранения энергии. Г. Гельмгольц и М.Рубнер продолжили исследования этого закона на живых организмах. Работами нем. учёных Г. Гельмгольца, Э. Дюбуа-Реймона, Д. Бернштейна и ряда др. были заложены основы представлений о механизме возникновения электрич. потенциалов в тканях и распространения возбуждения по нерву. Значение ионного состава и реакции среды в жизни клеток и тканей было выяснено в работах амер. исследователя Ж. Лёба, нем. учёных В. Нернста и Р. Гебера. В России И. М. Сеченов в конце 19 в. исследовал физич. закономерности растворения газов в крови и биомеханику движений. К. А. Тимирязев изучал фотосинтетич. активность отдельных участков солнечного спектра в связи с распределением энергии в нём и особенностями спектра поглощения хлорофилла (1903). А. Ф. Самойлов описал акустич. свойства среднего уха. П. П. Лазареву принадлежит заслуга в развитии ионной теории возбуждения (1916). М. Н. Шатер-ников использовал термодинамические представления в исследованиях энерге-тич. баланса организмов (1910-20). В 1905-15 были выполнены классич. исследования Н. К. Кольцова о роли физико-химич. факторов (поверхностного натяжения, концентрации водородных ионов, катионов) в жизни клетки. Этот этап предыстории Б., охватывающий период до 20гг. 20 в., характерен появлением отдельных работ с использованием идей и методов физики и физич. химии при исследовании движения, слухового и зрит, аппаратов, фотосинтеза, механизма генерации электродвижущей силы в нерве и мышце, значения ионной среды для жизнедеятельности клеток и тканей. После Октябрьской революции сложились благоприятные условия для развития Б. в СССР. В 1919 П. П. Лазарев создал в Москве Ин-т биологич. физики, где вели работы по ионной теории возбуждения, кинетике реакций, идущих под действием света, исследовали спектры поглощения и флуоресценции биологич. объектов, а также процессы первичного действия на организм различных факторов внеш. среды. Позже такие ин-ты были созданы и в др. странах. В 20-е гг. Кольцов сформулировал концепцию о молекулярной структуре гена и матричном механизме передачи наследств, информации и синтеза макромолекул. В 20-30-е гг. вышел ряд книг, оказавших глубокое влияние на последующее развитие Б. в СССР: "Биосфера" В. И. Вернадского (1926),"Теоретическая биология" Э. С. Бауэра (1935), "Физико-химические основы биологии" Д. Л. Рубинштейна (1932), "Организация клетки" Н. К. Кольцова (1936), "Реакция живого вещества на внешние воздействия" Д. Н. Насонова и В. Я. Александрова (1940). В эти годы шло постепенное формирование базы для биофизич. исследований, разрабатывались новые методы, росло технич. оснащение лабораторий. После 2-й мировой войны в СССР и ведущих капиталистич. странах в результате огромного размаха исследований по физике и химии, возникновения мощной приборостроительной пром-сти и резкого увеличения финансирования биологич. исследований начинается бурное развитие Б. Формирование отдельных областей Б. Молекулярная Б. исследует механизм биол. явлений с точки зрения взаимодействия атомов и молекул, ионов и радикалов. В задачу этого раздела входит изучение пространств, строения, физико-химич. свойств биологич. систем на молекулярном уровне. Эта проблематика тесно связана с биохимией, что особенно ярко видно на примере изучения строения биологически важных макромолекул, выяснение пространственной структуры к-рых требует биофизич. подхода и решается методом рентгеноструктурного анализа. Последний был успешно использован для расшифровки относительно простых биологич. молекул (в 20-х гг. в Англии В. Астбери удалось частично расшифровать структуру молекулы целлюлозы). Работы по структуре белка были начаты в 30-х гг. англ, учёным Дж. Берналом. К 1954 англ, исследователи Дж. Кендрю и М. Перу ц нашли метод расчёта пространственного расположения атомов в молекуле белка. Это позволило рассчитать структуру миоглобина и гемоглобина, что позволило вскрыть механизм возникновения серповиднокле-точной анемии и глубже понять природу активного центра белковой молекулы. Работы по изучению пространственной структуры белков ведутся в СССР на физич. ф-те МГУ, в Ин-те биофизики АН СССР и др. учреждениях. Исследования структуры фибриллярных белков (коллагена, фиброина шёлка) показали наличие регулярной структуры с периодически чередующимися группами аминокислот. Построена статистич. теория редупликации (удвоения) дезоксирибонуклеиновой кислоты (ДНК). К 1968 определена структура ок. 200 белков. Наряду с изучением строения отдельных молекул большие успехи достигнуты в исследовании молекулярных комплексов - ультраструктур, создающих функциональные единицы клетки. Исследования по молекулярной Б. тесно связаны с биохимией, генетикой и цитологией, молекулярной биологией. Значит, место в молекулярной Б. занимает проблема возбуждённых состояний молекул в биологич. системах; такие молекулы приобретают высокую химич. активность. Наиболее изучены возбуждённые состояния, возникающие на первичной стадии фотобиологич. процессов - фотосинтеза, зрения и биолюминесценции. Оригинальным направлением в отечеств. Б. можно считать изучение сверхслабого ультрафиолетового свечения биологич. систем (митогенетического излучения, А. Г. Гурвич, 1923-48). В 30-е гг. Г. М. Франк и С. Ф. Родионов разработали физич. метод обнаружения сверхслабых свечений биологич. объектов. Успехи в разработке методов регистрации сверхслабых световых потоков с помощью фотоэлектронных умножителей привели в 50-х гг. 20 в. к открытию сверхслабого свечения ряда животных и растит, объектов в видимой области спектра. Была показана связь этого свечения с рекомбинацией свободных радикалов. А. Н. Терениным с сотрудниками были исследованы механизмы элементарных фотофизич. процессов с участием пигментов, указана роль состояний молекул, открыт механизм миграции энергии в них при фотохимич. реакциях, изучен механизм люминесценции белков (1950-65). А. А. Красновский открыл и исследовал реакцию обратимого фотохимич. восстановления хлорофилла и его аналогов (1949-60). Эти работы способствовали развитию биологич. фотохимии. В одном из важных разделов Б. рассматривается превращение энергии в живых организмах, начиная с превращения и миграции энергии на молекулярном уровне и кончая энергетич. балансом целого организма (см. Биоэнергетика). Исследование взаимной трансформации химич. и механич. энергии при сокращении мышечного волокна, молекулярные механизмы движения ресничек и жгутиков у простейших, движения протоплазмы и клеточных органелл стали предметом изучения механохимии, находящейся на стыке биохимии и молекулярной Б. В 1938 в работе сов. учёных В. А. Энгельгардта и М. Н. Любимовой, изучавших механизм мышечного сокращения, было впервые продемонстрировано наличие прямой связи между механич. и химич. процессами. В дальнейшем эти работы были развиты амер. учёным А. Сент-Дьёрдьи. Традиционный раздел Б.- изучение физик о-х и ми ч. свойств клетки и проницаемости биологических мембран для различных веществ. Всё большее значение приобретают проблемы моделирования искусств, мембран и активного транспорта ионов. Одним из примеров практич. применения знаний, полученных в этой области ,Б., биохимией и физиологией, является создание искусственной почки. Важной проблемой Б. является изучение биоэлектрич. явлений. В этой области Б. тесно связана с физиологией (см. Биоэлектрические потенциалы). Исследования показали, что между наружной и внутр. средой каждой живой клетки поддерживается разность потенциалов ок. 0,1 в. Её источник - создаваемый клеткой ионный градиент между наружной и внутриклеточной средой. Эти данные послужили основой для создания мембранной теории генерации потенциалов в клетке, выдвинутой в начале века нем. учёным Д. Бернштейном и экспериментально обоснованной в 50-60-е гг. работами англ, учёных А. Ходжкина, А. Хаксли и Б. Каца, изучавших изменение проницаемости мембраны нервного волокна и ионные потоки в нерве при возбуждении (см. Мембранная теория возбуждения). Значит, место занимают также исследования др. физико-химич. свойств клеток - вязкости, оптических свойств, их изменений при различных физиологич. состояниях и тех или иных воздействиях. Биофизич. закономерности, свойственные организму в целом, рассматриваются в соответствующих разделах биоэнергетики (изучение механизма теплоотдачи, теплоизоляции, теплопродукции, скорости охлаждения при различных условиях и т. п.). Б. процессов управления неразрывно связана с кибернетикой биологической и биомеханикой. Созданию систем управления, выяснению принципов управления движениями животных и человека положили начало исследования сов. учёного Н. А. Бернштейна. Он первым приступил к изучению обратной связи в биологич. системах (1934). Изучение биомеханики движений (ходьба, бег, трудовые движения и др.), дыхания и кровообращения имеет исключит, важность в связи с вопросами физиологии труда и спорта, космическими полётами, а также для изучения причин сердечных и сосудистых заболеваний и создания аппаратов искусств, дыхания и кровообращения. Биофизич. исследования ведутся в СССР во мн. науч. учреждениях, в частности в Ин-те биофизики АН СССР, Ин-те цитологии АН СССР, Ин-те молекулярной биологии АН СССР, на кафедрах биофизики в МГУ, ЛГУ и в др. учреждениях. Одна из первых в мире кафедр Б. была осн. в МГУ в 1953 Б.Н.Тарусовым. Исследования по Б. и подготовка кадров ведутся во мн. странах мира. Великобритания - Лондонский ун-т, Ин-т молекулярной биологии, Кембридж; Венгрия - ун-т в г. Печ; Г Д Р-Ин-т биологии и медицины, Берлин; И з-р а и л ь - Ин-т Вейцмана, г. Реховот; Индия - Ин-т кристаллографии, молекулярной биологии и ядерной физики в Дели и ун-т в Мадрасе; К Н Р-Ин-т биофизики, Пекин; Польша - Варшавский ун-т и Ин-т биохимии и биофизики АН ПНР; Румыния - Ин-т биофизики, Бухарест; США - Йельский ун-т, Массачусетсский технологич. ин-т, Калифорнийский ун-т, Гарвардский ун-т, Рокфеллеровский ин-т и мн. др.; Франция - Ин-т физико-химич. биологии в Париже, Ин-т макромолекулярных исследований в Страсбуре и др.; Ф Р Г-Ин-т биофизики об-ва М. Планка, Франкфурт-на-Майне, Ин-т биологич. и мед. физики при Гёттингенском ун-те и др.; Чехословакия - Ин-т биофизики в Брно, Пражский ун-т; Швеция - Отделение биофизики при Нобелевском ин-те в Стокгольме; Япония - Ун-т в Осака, Ин-т белка, там же, Токийский ун-т. На 1-м Международном биофизич. конгрессе, состоявшемся в Стокгольме в 1961, был создан Международный союз теоретич. и прикладной биофизики, в центральный совет к-рого входят представители СССР. Периодические издания, в к-рых публикуются работы по Б.: "Биофизика" (М., 1956-); "Молекулярная биология" (М., 1967-); "Радиобиология" (М., 1961 -); "Advances in Biological and Medical Physics" (N. Y., 1948-); "Bio-chimica et Biophysica Acta" (N. Y.-Amst., 1947-); "Biophysical Journal" (N. Y., I960-); "Bulletin of Mathematical Biophysics" (Chi, 1939-); "Journal of Cell Biology" (N. Y., 1962-; в 1955-1961 наз.-"Journal of Biophysical and Biochemical Cytology"); "Journal of Molecular Biology" (N. Y.- L., 1959-); "Journal of Ultrastructure Research" (N. Y.- L., 1957-); "Progress in Biophysics and Biophysical Chemistry" (L., 1950-). Лит.: Бернштейн Н. А., О построении движений, М., 1947; Лазарев П. П., Сочинения, т. 2, М.- Л., 1950; Б р е с л е р С. Е., Введение в молекулярную биологию, М. -Л., 19б"6; Молекулярная биология. [Сб. ст.], пер. с англ., М., 1963; П а с ы н с к и й А. Г., Биофизическая химия, М., 1963; Аккерман Ю., Биофизика, пер. с англ., М., 1964; Вопросы биофизики. Материалы I Международного биофизического конгресса. Стокгольм, июль - август 1961, М., 1964; Сетлоу Р., Поллард Э., Молекулярная биофизика, пер. с англ., М., 1964; Волькенштейн М. В., Молекулы и жизнь. Введение в молекулярную биофизику, М., 1965; Биофизика, М., 1968; Саsеу Е., Biophysics. Concepts and mechanisms, N. Y. -L., 1962; Physical techniques in biological research, v. i -5, N. Y., 1955-64. Б. Н. Вепринцев. БИОФИЛЬТР, сооружение для биологич. очистки сточных вод. Представляет собой круглый или прямоугольный в плане резервуар с двойным дном, наполненный фильтрующим материалом (котельный шлак, гранитный щебень, гравий, керамзит и др.). Расстояние между днищами не менее 0,4 м. Высота фильтрующего слоя 1,5-2 м; крупность зёрен фильтрующего слоя 30-50 мм, нижнего подстилающего слоя - 60-100 мм. При прохождении сточной воды через фильтрующий материал на его поверхности образуется биологич. плёнка из скоплений бактерий, грибков, окисляющих и минерализующих органич. вещества сточной воды. Окислительная мощность Б. определяется опытным путём. См. Биологическая очистка сточных вод. БИОХИМИИ ИНСТИТУТ им. А. Н. Баха АН СССР. Организован в Москве в 1935 по инициативе А. Н. Баха и А. И. Опарина. Основная задача Б. и. - изучение процессов обмена веществ и энергии в живых организмах и способов управления процессами обмена. В числе важнейших проблем, разрабатываемых Б. и.: биология ферментов, механизм их действия в живой клетке и поведение в автолитич. смесях; биологич. фиксация азота; обмен аминокислот у растений и микроорганизмов; биологич. окисление; роль фосфорных соединений в дыхательных процессах; природа первичных процессов, происходящих при фотосинтезе в растениях и бактериях; биохимия фитоиммунитета; биосинтез и механизм действия витаминов; химия и биохимия углеводов; эволюционная биохимия и радиобиология. Важное значение имеют работы А.И. Опарина по проблеме возникновения жизни на Земле. Исследования, выполненные в Б. и., нашли применение в народном хозяйстве и способствовали усовершенствованию ряда старых и возникновению новых отраслей пром-сти. Напр., выяснена био-химич. сущность изменений, происходящих в процессе хранения зерна и его переработки. Предложены и внедрены в промышленность способы получения препаратов витаминов A, Bi2 для мед. целей и для кормовых концентратов; разработан и внедрён в произ-во микробиологич. метод получения L-лизина в кристаллич. виде и в виде кормового концентрата, содержащего 15-18% лизина, и др. К началу 1970 в Б. и. было 14 лабораторий. Труды сотрудников Б. и. публикуются преим. в журналах: "Биохимия", "Доклады АН СССР", "Известия АН СССР", "Прикладная биохимия и микробиология", в тематич. сборниках и в виде монографий. Б. и. имеет аспирантуру. В 1967 награждён орденом Ленина. Н. Н. Дьячков. БИОХИМИЧЕСКАЯ ОЧИСТКА сточных вод, см. Биологическая очистка сточных вод. БИОХИМИЧЕСКОЕ ОБЩЕСТВО Всесоюзное (ВБО), научное общество, объединяющее советских учёных, работающих в области биохимии. Организовано в 1958 при АН СССР. Отделения об-ва имеются в союзных и автономных республиках, краях и городах СССР. В 1959 насчитывалось 1570 членов об-ва, в 1964 было 3500, а в 1970 в его состав входило 15 республиканских об-в и 53 городских отделения, объединивших св. 6500 биохимиков. Основные задачи об-ва: содействие развитию в СССР всех отраслей биохимии, оказание членам об-ва помощи в повышении квалификации, науч. и методич. помощи в постановке преподавания биохимии в высшей и средней школе; популяризация и пропаганда знаний и новейших научных и практич. достижений в биохимии, содействие развитию науч. связей с зарубежными учёными и т. д. Общество проводит конгрессы, съезды, конференции по различным разделам биохимии; циклы лекций, доклады, семинары для повышения квалификации своих членов, осуществляет науч. связи с др. науч. об-вами, издаёт ежегодник "Успехи биологической химии" (с 1950), труды биохимич. съездов, конференций, симпозиумов; ВБО - член Междунар. биохимич. союза, а также федерации Европ. биохимич. об-в. В 1959 на делегатском съезде ВБО был принят устав об-ва и избран Центральный совет. Президенты об-ва -А. И. Опарин (1958-64), А. В. Палладии (1964-69), С. Е. Северин (с 1969). Лит.: Будницкая Е. В., Во Всесоюзном биохимическом обществе, "Биохимия", 1959, т. 24, в. 3; 1964, т. 29, в. 4; Первый Всесоюзный биохимический съезд, там же, 1966, т. 31, в. 3. А. А. Лизандр. БИОХИМИЯ, биологическая химия, наука, изучающая состав организмов, структуру, свойства и локализацию обнаруживаемых в них соединений, пути и закономерности их образования, последовательность и механизмы превращений, а также их биологич. и физиоло-гич. роль. В зависимости от объекта исследования Б. подразделяют на Б. микробов, растений, животных и человека. Это подразделение условно, т. к. в составе различных объектов и в протекающих в них биохимич. процессах много общего. Поэтому результаты исследований, проведённых на микробах, растит, или животных тканях и клетках, взаимно дополняют и обогащают друг друга. Тесно связаны между собой и разные направления биохимических исследований, однако принято делить Б. на статическую, занимающуюся преим. анализом состава организмов, динамическую, изучающую превращения веществ, и функциональную, выясняющую, какие химич. процессы лежат в основе различных проявлений жизнедеятельности. Это последнее направление исследований иногда выделяют под спец. назв. физиологическая химия. Вся совокупность химич. реакций, протекающих в организмах, включая усвоение веществ, поступающих извне (ассимиляция), и их расщепление (диссимиляция) вплоть до образования конечных продуктов, подлежащих выделению, составляет сущность и содержание обмена веществ -главного и постоянного признака всего живого. Понятно, что изучение обмена веществ во всех деталях - одна из осн. задач Б. Биохимич. исследования охватывают очень широкий круг вопросов: нет такой отрасли теоретич. или прикладной биологии, химии и медицины, к-рая не была бы связана с Б., поэтому совр. Б. объединяет ряд смежных научных дисциплин, ставших с середины 20 в. самостоятельными. Накопление биохимич. сведений и формирование Б. в 16-19 вв. Б. сформировалась как самостоятельная наука в конце 19 в., хотя истоки её относятся к далёкому прошлому. С 1-й пол. 16 в. и до 2-й пол. 17 в. свой вклад в развитие химии и медицины вносили ятрохимики (химики-врачи): нем. врач и естествоиспытатель Ф. Парацельс, голл. учёные Я. Б. ван Гельмонт, Ф. Сильвий и др., занимавшиеся исследованием пищеварит. соков, жёлчи, а также процессов брожения (см. Ятрохимия). Сильвий, прославленный врач, придавал особенно большое значение правильному соотношению в организме человека кислот и щелочей; он полагал, что в основе многих, если не всех, болезней лежит расстройство этого соотношения. Большая часть принятых ятрохимиками положений была наивной, полной заблуждений; однако нельзя забывать, что науч. химии тогда ещё не существовало. Наиболее общей теорией, господствовавшей в науке того времени, была теория т. н. флогистона. Тем не менее балансовые опыты на человеке с точным учётом массы тела и выделений были проведены итал. учёным С. Санторио в начале 17 в. Эти опыты привели к описанию "perspiratio insensibilis" - потери массы за счёт чнечувствуемого пропотевания". Великие открытия в области физики и химии 18 и нач. 19 вв. (открытие ряда простых веществ и соединений, формулировка газовых законов, открытие законов сохранения материи и энергии) заложили науч. фундамент общей химии. После открытия в составе воздуха кислорода голл. ботаник Я. Ингенхауз смог описать постоянное образование растением СО2 и выделение на солнечном свету зелёными частями растения кислорода. Опытами Ингенхауза было положено начало исследованию дыхания растений и процессов фотосинтеза, детальное изучение к-рых продолжается и в настоящее время. В кон. 1-й четв. 19 в. было известно очень ограниченное кол-во органич. веществ. В учебнике нем. химика Л. Гме-лина, изданном в 1822, упоминается лишь 80 органич. соединений. Задачи и возможности органич. химии в то время оставались неясными. Швед, учёный И. Берцелиус считал, что органические тела разделяются на два чётко разграниченных класса - на растения и животные; сущность живого тела основана не на его неорганич. элементах, а на чём-то ином. Это нечто, что он называет "жизненной силой", лежит целиком за пределами неорганических элементов. Берце-лиус выражает сомнение в том, что люди когда-либо сумеют искусственно производить органические вещества и подтвердить анализ синтезом (1827). Несостоятельность таких типичных для витализма позиций выявилась очень скоро. Уже в 1828 нем. химик Ф. Вёлер, ученик Берцелиуса, получил синтетич. путём мочевину, описанную в 18 в. франц. учёным Г. Руэлем в качестве составной части мочи млекопитающих. Вскоре последовали синтезы др. как природных органич. соединений, так и искусств., неизвестных в природе. Т. о. рушилась стена, отделявшая органич. соединения от неорганических. Начиная со 2-й пол. 19 в. органич. химия становится всё больше химией синтетической, усилия к-рой направляются на получение новых соединений углерода, особенно имеющих пром. значение; в её задачи уже не входит исследование состава растит, и животных объектов. Эти сведения поступали случайно в результате побочной работы химиков, ботаников, физиологов растений и животных, а также патологов и врачей, включавших в круг своих интересов химич. исследования. Так, в 1814 рус. химик К. С. Кирхгоф описал осахаривание крахмала под влиянием вытяжки из проросших семян ячменя: действие амилазы. К сер. 19 в. были описаны и др. ферменты: амилаза слюны, расщепляющая полисахариды; пепсин желудочного сока и трипсин сока поджелудочной железы, расщепляющие белки. Берцелиус ввёл в химию понятие катализаторов, к числу к-рых были отнесены все известные в то время ферменты. В 1835 франц. химик М. Шеврёль описал в составе мышц креатин, несколько позднее в моче был найден близкий к нему по структуре креатинин. Содержание в скелетных мышцах молочной кислоты и её накопление при работе установил нем. химик Ю. Либих. В 1839 он же выяснил, что в состав пиши входят белки, жиры и углеводы, являющиеся главными составными частями животных и растит, организмов. В сер. 19 в. была установлена структура жира и осуществлён его синтез франц. химиком П. Бертло; синтез углеводов был проведён рус. учёным А. М. Бутлеровым; он же предложил теорию строения органических соединений, сохранившую своё значение и поныне. Систематич. исследование белков было начато голл. врачом и химиком Г. И. Мульдером в 30-е гг. 19 в. и интенсивно продолжалось многими авторами во все последующие годы. В то же время в связи с описанием дрожжевых клеток (К. Конь-яр-Латур во Франции и Т. Шванн в Германии, 1836-38) активно начали изучать процесс сбраживания сахара и образования спирта, издавна привлекавший к себе внимание. В числе учёных, изучавших брожение, были Ю. Либих и франц. учёный Л. Пастер. Пастер пришёл к выводу, что брожение - биологич. процесс, в к-ром обязательно участвуют живые дрожжевые клетки. Либих же рассматривал сбраживание сахара как сложную химич. реакцию. В этот спор была внесена ясность, когда рус. химик М. М. Манассеина (1871) и особенно чётко нем. учёный Э. Бухнер (1897) доказали способность бесклеточного дрожжевого сока вызывать алкогольное брожение. Т. о. была подтверждена принципиальная правильность хим. теории действия ферментов, к-рую Либих сформулировал в 1870; осн. принципы этой теории сохранили своё значение и теперь. Постепенно количество накопившихся сведений относительно хим. состава растит, и животных организмов и протекающих в них хим. реакций стало значительным, в связи с чем были осуществлены попытки их систематизации и объединения в учебных руководствах. Наиболее ранние из них - учебники И. Зимона (1842) и Либиха (1847), изданные в Германии, и учебник физиологич. химии А. И. Ходнева, вышедший в России (1847). Возникновение и развитие современных направлений Б. В конце 19 века и в 20 в. развитие Б. приобрело выраженный специализированный характер в зависимости от разрабатываемой проблемы и объекта исследования. Б. растений развивалась по преимуществу на кафедрах ботаники и физиологии растений. Тесно связана с ней и Б. микроорганизмов. Белки, углеводы, ли-пиды, витамины, являющиеся составными частями растений, животных и микроорганизмов, исследовали биохимики всех стран на самых различных объектах. Характерными для растений и микроорганизмов можно считать гликозиды, дубильные вещества, эфирные масла, алкалоиды, антибиотики и др. т. н. вещества вторичного происхождения. Из перечисленных соединений ряд гликозидов был синтезирован при участии ферментов франц. химиком Э. Буркло и его сотрудниками (1911-18). В расшифровке строения антоцианов - гликозидов, входящих в состав пигментов цветов и плодов, - исключит, роль сыграли классич. работы нем. химика Р. Вильштеттера (1910-15). Группа алкалоидов (азотистых гетероциклич. веществ основного характера) изучалась нем. химиком А. Гофманом (1890-1900). Позднее алкалоиды изучали выдающиеся исследователи (Р. Вилынтеттер, А. Пикте - Швейцария; рус. химики А. П. Орехов, А. А. Шмук и мн. др.). Эфирные масла, терпены успешно исследовали также крупные представители химии и биохимии: Перкин мл. (Великобритания), Г. Эйлер (Швеция) и др. Выдающуюся роль в развитии Б. растений в России (конец 19 в. - 1-я половина 20 в.) сыграли проф. Петерб. ун-та А. С. Фаминцын, его ученики Д. И. Ивановский, открывший вирусы, и И. П. Бородин, изучавший окислит, процессы в организме растений и их связь с превращениями белков. Работы С. П. Костычева (проф. Петербургского университета, позднее -ЛГУ) по анаэробному обмену углеводов и дыханию у растений обогатили хим. физиологию открытием новых промежуточных продуктов брожения, формулировкой оригинальных взглядов на сущность окислительных процессов, на обмен белков и фиксацию азота растениями. Много сделал проф. Варшавского ун-та М. С. Цвет, разработавший метод хроматографии на колонках, используемый и в наст, время. Моск. школа физиологов и биохимиков растений была представлена К. А. Тимирязевым, исследовавшим фотосинтез и химию хлорофилла. Его ученики - В. И. Палладии, разрабатывавший проблему биологич. окисления, Д. П. Прянишников, изучавший азотистый обмен растений, В. С. Буткевич, обогативший теоретич. Б. исследованиями белков и белкового обмена растений, А. Р. Кизель, изучавший обмен аргинина и мочевины у растений и структурные элементы протоплазмы клеток,- явились создателями крупных школ и оригинальных направлений совр. общей и эволюционной Б., а также физиологии и Б. растений, плодотворно развивающихся и в 3-й четв. 20 в. В 20 в. представители Б. микроорганизмов и Б. растений решали много общих задач, связанных с изучением природных соединений (в т. ч. и высокомолекулярных), их структуры, путей образования и расщепления, характеристики ферментов, участвующих в этих процессах. Следует отметить, что микроорганизмы постепенно стали излюбленным объектом для различных энзимологич. исследований и для разработки проблем биохим. генетики. Все эти исследования создали прочную базу для разработки мн. частных проблем, в т. ч. и пром. Б. К ним относятся получение новых антибиотиков, разработка методов их очистки, поиски условий, благоприятных для микробиол. синтеза не только антибиотиков, но и др. биологически активных соединений - витаминов, дефицитных аминокислот, нуклео-тидов и т. д. Техническая и промышленная Б. Потребности нар. х-ва -проблемы рентабельного получения сырья, его удобного и рационального хранения, правильной обработки и эффективного использования, а также проблемы повышения урожайности культурных растений, вопросы виноградарства и технологии виноделия, запросы пищ. пром-сти - привели к созданию новых отраслей Б.- технич. и пром. Б. В СССР это направление представлено наиболее полновесно в Ин-те биохимии им. А. Н. Баха (А. И. Опарин, В. Л. Кретович, Л. В. Метлицкий, Р. М. Фениксова и др.), в Ин-те физиологии растений АН СССР (А. Л. Курсанов, его сотрудники и ученики). Много сделали в изучении биохимии зерновых культур И. П. Иванов (Всесоюзный ин-т растениеводства), а также В. Л. Кретович, М. И. Княгиничев, их сотрудники и мн. др. Работы, проведённые в Ин-те им. А. Н. Баха по Б. катехинов, сыграли существенную роль в развитии чайного произ-ва и дубильных веществ. Б. животных и человека (мед. и физиологич. химия). Большое значение для развития этой ветви Б. имели многочисленные школы физиологов, химиков, патологов и врачей, работавших в разных странах. Во Франции в лаборатории физиолога К. Бернара в составе печени млекопитающих был открыт гликоген (1857), изучены пути его образования имеханизмы, регулирующие его расщепление; здесь же Л. Корвизар (1856) открыл в поджелудочном соке фермент трипсин. В Германии в лабораториях Ф. Хоппе-Зейлера, А. Косселя, Э. Фишера, Э. Абдергальдена, О. Хаммарстена и др. подробно изучались простые и сложные белки, их структура и свойства, вещества, образующиеся при искусственном их расщеплении путём нагревания с кислотами и щелочами, а также под влиянием ферментов. В Англии Ф. Хопкинс, основатель школы биохимиков в Кембридже, занимался исследованием аминокислотного состава белков, открыл триптофан, глутатион, изучал роль аминокислот и витаминов в питании. Существенный вклад в развитие Б. в кон. 19 - нач. 20 вв. внесли русские учёные, работавшие на кафедрах высших учебных заведений и в специализированных институтах. В Военно-мед. академии А. Я. Данилевский и его сотрудники разрабатывали проблемы химии белка, методы выделения и очистки ферментов, изучали механизм их действия и условия обратимости ферментативных реакций. В Ин-те экспериментальной медицины М. В. Ненцкий исследовал химию порфиринов, биосинтез мочевины, а также ферменты бактерий, вызывающие разложение аминокислот. Особенно плодотворным было содружество лабораторий А. Я. Данилевского и М. В. Ненцкого с лабораторией И. П. Павлова при исследовании пищеварения и образования мочевины в печени. В Моск. ун-те В. С. Гулевич подробно и успешно исследовал азотистые экстрактивные (небелковые) вещества мышц и открыл ряд новых соединений оригинальной структуры (карнозин, карнитин и др.). Предметом многочисленных исследований было и остаётся подробное изучение разнообразных ферментативных реакций, протекающих в паренхиматозных органах, гл. обр. в печени, и обусловливающих нормальное течение процессов обмена веществ. Большое внимание во 2-й пол. 19 и в 20 вв. было уделено биохим. исследованию возбудимых тканей, гл. образом мозга и мышц. В СССР разработка этих проблем осуществлялась А. В. Палладиным, Г. Е. Владимировым, Е. М'. Крепсом, их учениками и сотрудниками. К середине 20 в. нейрохимия представляла одно из сформировавшихся самостоятельных направлений Подверглась всестороннему изучению Б. крови. Дыхательная функция крови (т. е. связывание и отдача кровью углекислого газа и кислорода), изучавшаяся в середине 19 в. в лаборатории К. Людвига в Вене, подробно исследовалась в дальнейшем в разных странах. Полученные данные привели к анализу структуры и свойств гемоглобина в норме и патологии, к детальному изучению реакции между гемоглобином и кислородом и выяснению закономерностей кислотно-щелочного равновесия. Крупных успехов Б. достигла в изучении витаминов, гормонов, минеральных веществ, в частности микроэлементов, их распространения в различных организмах, физиологич. роли, механизма действия и регулирующих влияний на ферментативные реакции и процессы обмена веществ. Большое значение имеет проблема связи структуры и функции, к-рая характеризует также задачи биохимич. фармакологии, когда речь идёт о лекарств, средствах и исследовании первичного механизма их действия, осуществляемого вмешательством в ферментативные реакции, составляющие основу процессов обмена веществ. В сер. 20 в. самостоят, значение приобрели биохим. исследования, проводившиеся в клиниках и посвящённые изучению биохимич. особенностей организма, химич. состава крови, мочи и др. жидкостей и тканей больного человека. Это направление, получившее широкое развитие, составляет осн. содержание клинической Б. Витаминология. В лаборатории Г. А. Бунге молодой рус. врач Н. И. Лунин первый описал в 1880 в составе молока добавочные факторы питания. В 1896 аналогичное наблюдение было сделано голл. врачом К. Эйкманом, описавшим присутствие важного для организма фактора в рисовых отрубях. Польский исследователь К. Функ в 1912 выделил активное начало в кристаллич. виде и назвал его витамином. Работы этого направления получили широкое развитие; постепенно были открыты мн. др. витамины, и сейчас витаминология представляет один из весьма важных разделов Б., а также науки о питании. Б. гормонов. Работы, связанные с анализом химич. структуры продуктов жизнедеятельности желез внутренней секреции - гормонов, путей их образования в организме, механизма действия и возможного осуществления лабораторного синтеза, представляют одно из важных направлений биохимич. исследований. Б. стероидных гормонов - часть общей проблемы Б. стеринов. Достигнутые в этой области успехи в значительной мере связаны с использованием меченных по углероду (С14) исходных и промежуточных соединений. Самая тесная связь установилась между широким фронтом исследований белковых веществ и специальным изучением структуры и функций гормонов белковой природы. Изучение гормональной активности тех или других препаратов невозможно без глубокого анализа биохимич. механизма их действия. Т. о., данные по химии и Б. гормонов в равной мере обогащают эндокринологию и Б. Энзимологияи - учение о ферментах, вполне самостоятельная область Б. В ней проблема строения белков-ферментов тесно переплетается с физико-химич. проблемами - химич. кинетикой и катализом. В 3-й четв. 20 в. внесено много нового в представления о структуре ферментов, о их присутствии в нативном состоянии в виде сложных комплексов. Анализ строения ферментов в сопоставлении с проявляемой ими в разных условиях активностью позволил выяснить значение отдельных аминокислот (гл. обр. цистеина, лизина, гистидина, тирозина, серина и т. д.) в формировании активного центра ферментов. Выяснены структура мн. коферментов, их значение для ферментативной активности, а также связь между коферментами и витаминами. Большой вклад в развитие энзимологии в первой половине 20 в. внесли Р. Вилыптеттер, Л. Михаэлис, Г. Эмбден, О. Мейергоф (Германия), Дж. Самнер, Дж. Нортроп (США), Г. Эйлер (Швеция), А. Н. Бах (СССР). Много сделали продолжающие активно работать создатели крупных школ и направлений: О.Варбург (Зап. Берлин), Ф. Линен (ФРГ), Р. Питере, X. Кребс (Великобритания), X. Теорелль (Швеция), Ф. Липман, Д. Кошленд (США), А. Россн-Фанелли (Италия), Ф. Шорм (Чехословакия), Ф. Штрауб (Венгрия), Т. Барановский, Ю. Хеллер (Польша) и мн. др. В СССР эту область исследований представляют: В. А. Энгельгардт и М. Н. Любимова, установившие ферментативную активность мышечных белков, в частности аденозинтрифосфатазную активность миозина и процесс окислительного фосфорилирования, А.Е. Браунштейн, открывший совм. с М. Г. Крицман процесс переноса аминогруппы (переаминирование); А. И. Опарин и А. Л. Курсанов, изучавшие роль структуры клеток в проявлении активности ферментов; С. Р. Мардашев, успешно исследовавший декарбоксилирование аминокислот, и др. Исследования сложных комплексов ферментов проводятся в лабораториях Л. Рида (США), М. Койке (Япония), В. Санади (США), Ф. Линена (ФРГ), С. Е. Северина (СССР) и др. Сов. учёный В. А. Белицер значительно углубил представления об энергетической эффективности открытого В. А. Энгельгардтом дыхательного пути образования богатых энергией соединений; Г. Е. Владимиров уточнил количество энергии (Юкал, или 42 дж), освобождающееся при гидролизе АТФ (см. Аденозинфосфорные кислоты). Работы этого направления, сначала остававшиеся единичными, в 50-е и последующие годы получили очень широкое развитие гл. обр. в результате исследований Д. Грина и Б. Чанса, А. Ленинджера, Э. Рэккера (США), Э. Слатера (Нидерланды), Л. Эрнстера (Швеция) и др. В СССР эта проблема разрабатывалась в МГУ и ЛГУ на кафедрах Б., а также в отдельных лабораториях (С. А. Нейфах, В. П. Скулачев и др.). Современные исследования показали также наличие выраженного влияния солевого состава среды и отдельных ионов на ферментативные процессы и важную роль микроэлементов в реализации ферментативной активности. Эволюционная и сравнительная Б. Исследования по Б. животных, растений и микроорганизмов показали, что, несмотря на общность осн. биохимич. структур и процессов у всех живых организмов, имеются и специфич. различия, зависящие от уровня онто- и филогенетич. развития изучавшихся объектов. Накопленные факты позволили заложить фундамент сравнительной Б., задача к-рой - найти закономерности биохимич. эволюции организмов. Большое теоретич. значение имеет проблема происхождения жизни на Земле. Нек-рые важные положения теории А. И. Опарина о происхождении жизни получили экспериментальное подтверждение в работах Ин-та им. Баха, кафедры Б. растений МГУ и ряда зарубежных лабораторий (И. Оро, С. У. Фоке в США; и др.). Гистохимия. Цитохимия. По мере развития техники морфологич. исследований, особенно после введения в практику лабораторной работы электронной микроскопии, открывшей много-числ., ранее неизвестные структуры в составе клеточного ядра и протоплазмы, перед Б. встали новые задачи. На стыке морфологич. л биохимич. исследований возникли новые отрасли - гистохимия и цитохимия, изучающие локализацию и превращение веществ в клетках и тканях и использующие биохимич. и морфологич. методы. Биоорганическая химия. Подробные исследования структуры биополимеров - простых и сложных белков, нуклеиновых кислот, полисахари-дов и липидов, а также анализ действия биологически активных низкомолекулярных природных соединений (коферментов, нуклеотидов, витаминов и т. д.) привели к необходимости изучения связи между строением вещества и его биологич. функцией. Постановка этого вопроса вызвала развитие исследований, находящихся на грани биологич. и органич. химии. Данное направление исследований получило наименование биоорганической химии. Молекулярная биология. Разработка методов разделения субклеточных структур (ультрацентрифугирование) и получение отдельно фракций, содержащих клеточные ядра, митохондрии, рибосомы. и т. п., позволили детально исследовать состав и биологич. функции выделенных образований. Применение методов электрофореза в сочетании с хроматографией дало возможность детально характеризовать высокомолекулярные соединения. Параллельно улучшалась техника аналитич. определений, позволявшая исследовать ничтожное количество материала. Это было связано с внедрением в биологию, в т. ч. и в Б., физических (гл. обр. оптических) методов исследования (флуорометрия, спектрофотомет-рия в различных областях спектра, масс-спектрометрия, ядерномагнитный и электронно-парамагнитный резонанс, газово-жидкостная хроматография), с применением радиоактивных изотопов, чувствительных автоматич. анализаторов аминокислот, пептидов, нуклеотидов, поляро-графии, высоковольтного электрофореза и т. д. Всё это привело к появлению ещё одного самостоятельного ответвления Б., тесно связанного с биофизикой и физической химией и названного молекулярной биологией. Составной частью молекулярной биологии можно считать молекулярную генетику, несмотря на нек-рые специфич. её задачи. Так, напр., анализ механизма возникновения ряда наследственных нарушений обмена веществ и функций организма позволил выяснить роль выпадения или извращения биосинтеза тех или иных белковых веществ, обладающих ферментативной, иммунной или др. биологич. активностью. Сюда относятся также исследования нарушений в обмене углеводов, аминокислот (напр., фенилаланина, тирозина, триптофана и др.), образования пато-логич. форм гемоглобина и т. д. Благодаря развитию новых методов исследования Б. в 1950-1970 гг. достигла крупных успехов. Это прежде всего -выяснение строения белков, определение последовательности расположения в них аминокислот. Впервые была выяснена последовательность расположения аминокислот в гормоне белковой природы -инсулине - англ, биохимиком Ф. Сангером, затем в ферменте рибонуклеазе К.Хёрсом, С. Муром и У.Стейном (США), разработавшими метод автоматич. анализа аминокислот, вошедший в практику биохимич. лабораторий. Тот же фермент - рибонуклеазу, полученную из разных источников, изучали К. Анфинсен (США), Ф. Эгами (Япония) и др. Последовательность расположения аминокислот в ряде протеолитических ферментов установили Ф. Шорм и Б. Кейль с сотрудниками (Чехословакия), Б. Хартли (Великобритания) и др. Большое достижение Б. 60-х гг. 20 в. - химич. синтез гормонов - адренокортикотропного гормона, молекула которого содержит 23 аминокислоты (в природном гормоне 39 аминокислот), и инсулина, молекула к-рого состоит из 51 аминокислоты, фермента рибонуклеазы (124 аминокислоты). В СССР над проблемами структуры и синтеза биологически активных веществ работают в Ин-те химии природных соединений (директор М. М. Шемякин), Ин-те биологич. и мед. химии (директор В. Н. Орехович) и др. ин-тах и на кафедрах вузов. С большим успехом использовали англ, учёные М. Перуц, Дж. Кендрю и их сотрудники рентгеноструктурный анализ для выяснения строения миоглобина и гемоглобина. В 1956-67 полностью была определена структура лизоцима англ. биохимиком Д. Филлипсом и др. Не менее значительны успехи, достигнутые в анализе сложных белков, нуклеопротеидов, нуклеиновых кислот и нуклеотидов. Триумфом Б., молекулярной биология и генетики явились исследования, показавшие роль нуклеиновых кислот в биосинтезе белков и установившие предопределяющее влияние нуклеиновых кислот на строение и свойства синтезируемых в клетке белков. Этими работами были выяснены биохимич. основы передачи признаков по наследству от поколения к поколению. Трудно переоценить также значение исследований, определивших последовательность нуклеотидов в составе транспортных рибонуклеиновых кислот и разработку методов органич. синтеза полинуклеотидов. Особенно плодотворно в названных областях работают И. Бьюкенен, Э. Чаргафф, И. Дэвидсон, Д.Дейвис, А. Корнберг, С. Очоа, Дж. Уотсон, М. Уилкинс и др. (США), Ф. Крик, Ф. Сангер (Великобритания), Ф. Жакоб, Ж. Моно (Франция), А. Н. Белозерский, А. С. Спирин, В. А. Энгельгардт, А. А. Баев (СССР) и мн. др. Каучные учреждения, общества и периодич. издания. Запросы к Б. со стороны смежных науч. дисциплин - медицины со всеми её разветвлениями, с. х-ва (растениеводства, животноводства), пищ. пром-сти, теоретич. и прикладной биилогии, почвоведения, гидробиологии и океанологии, становятся всё шире. Каждое из направлений Б. располагает в СССР и за рубежом сетью специализированных ин-тов и лабораторий. Науч. работа по Б. в СССР проводится в центральных н.-и. ин-тах, находящихся в системе: АН СССР - Ин-т биохимии им. А. Н. Боха, Ин-т эволюционной физиологии и биохимии, Ин-т физиологии ранений, Ин-т молекулярной биологии, Ин-т химия природных соединений; республиканских академий - ин-ты биохимии УССР, Ар;.. ССР, Узб. ССР, Лптов. ССР; отраслевых академий: Ин-т биологич. и мед. химии АМН СССР, отдел биохимии в Ин-те эксперимент, медицины АМН СССР, Ин-т эксперимент, эндокринологии и химии гормонов АМН СССР, Ин-т питания АМН СССР; в ин-тах ВАСХНИЛ и ряда министерств (здравоохранения, с. х-ва, пищевой пром-сти и др.). Работы по Б. представлены в лаборатории биоорганической химии МГУ, на многочисл. кафедрах Б. вузов. Проблемами Б. занимаются в центральных и отраслевых институтах, работающих в области ботаники, физиологии, патологии, в ин-тах экспериментальной и клинич. медицины, ин-тах пищ. пром-сти, ин-тах физкультуры и мн. др. Осн. специалистов-биохимиков за рубежом и в СССР готовят ун-ты, их химич. и биологич. ф-ты, имеющие в своём составе специальные кафедры. Биохимиков более узкого профиля готовят в медицинских, технологич., с.-х. и др. вузах. В большинстве стран существуют науч. биохимические общества, объединённые в Европейскую федерацию биохимиков (FEBS - Federation of European Biochemical Societies) и в Междунар. биохимич. союз (IUB - International Union of Biochemistry). Эти организации собирают симпозиумы, конференции, а также конгрессы - ежегодные по Европ. федерации (первый проходил в 1964) и раз в 3 года по Междунар. биохимич. союзу (первый состоялся в 1949; особенно популярными и многолюдными конгрессы стали начиная с 5-го, состоявшегося в 1961 в Москве). В СССР Всесоюзное биохимич. об-во с многочисленными республиканскими и городскими отделениями было организовано в 1958. Оно объединяет ок. 6,5 тыс. членов. Фактически число биохимиков в СССР значительно больше. Количество периодич. изданий, в к-рых публикуются работы по Б., очень велико и продолжает увеличиваться с каждым годом. Из зарубежных и международных журналов наиболее известны: "Journal of Biological Chemistry" (Bait., 1905-), "Biochemistry" (Wash., 1964-), "Archives of Biochemistry and Biophysics" (N. Y., 1942-), "Biochemical Journal" (L., 1906-), "Phytochemistry" (Oxf.-N. Y., 1962-), "Molecular Biology" (издаётся в Англии - журнал международный), "Bulletin de la Societe de Chimie Biologique" (P., 1914-), "Enzymologia" (Haaga, 1936-), "Giornale di Biochimica" (Rome, 1955-),"Acta Biological et Medica Germanica"(Lpz., 1959-), "Hoppe Seyler's Zeitschrift fur physiologische Chemie" (Berlin, 1877-)," Journal of Biochemistry" (Tokyo, 1922-). Популярны ежегодники: "Annual Review of Biochemistry" (Stanford, 1932-), "Advances in Enzymology and Related Subjects of Biochemistry" (N. Y., 1945-), "Advances in Protein Chemistry" (N. Y., 1945-), "Advances in Enzyme Regulation" (Oxf., 1963-), "Advances in Molecular Biology" и др. В СССР экспериментальные работы по Б. печатаются в журналах: "Биохимия" (М., 1936-), "Журнал эволюционной биохимии и физиологии" (М., 1965-), "Молекулярная биология" (М., 1967-), "Вопросы медицинской химии" (М., 1955-), "Украинский биохимический журнал" (К., 1926-), "Прикладная биохимия и микробиология" (М., 1965-), "Доклады АН СССР" (М., 1933-), "Бюллетень экспериментальной биологии и медицины" (М., 1936-), "Известия АН СССР. Серия биологии и медицины" (М., 1936-), "Известия АН СССР. Серия химическая" (М., 1936-), "Научные доклады высшей школы. Серия биологические науки" (М., 1958-) и в некоторых др. Обзорные работы по Б. печатаются в журнале "Успехи современной биологии" (М., 1932-), в ежегоднике "Успехи биологической химии" (т. 1-8, 1950-67), издаваемом Всесоюзным биохимическим обществом, в журн. "Успехи химии" (М., 1932-), "Реферативный журнал. Химия. Биологическая химия" (М., 1955-), в журн. Всесоюзного общества им. Менделеева. Часто выходят в свет труды биохимич. ин-тов. Лит.: Руководства: Макеев И. А., Гулевич В. С., Б р о у д е Л. М., Курс биологической химии, М., 1947; К р е т о в и ч В. Л., Основы биохимии растений, 4 изд., М., 1964; 3 б а р с к и й Б. И., Иванов И. И., М а р д а ш е в С. Р., Биологическая химия, 4 изд., М., 1965; Фердман Д. Л., Биохимия, 3 изд., М., 1966. История: Прянишников Д., Избр. соч., т. 1, М., 1951, с. 5 -19; Гулевич В. С., Избранные труды, М., 1954, с. 5-21; Парнас Я. О., Избранные труды, М., 1960, с. 5 - 10; Толкачевская Н. Ф., Развитие биохимии животных, М., 1963; Джуа М., История химии, пер. с итал., М., 1966; Развитие биологии в СССР, М., 1967; Кретович В. Л., Введение в энзимологию, М., 1967; Биохимия растений, пер. с англ., М., 1968; Lieben F., Geschichte der physiologischen Chemie, Lpz.- W., 1935. Монографии: Энгельгардт В. А., Некоторые проблемы современной биохимии, М., 1959; его же, Пути химии в познании явлений жизни, М., 1965; Северин С. Е., Биохимические основы жизни, М., 1961; С п и р и н А. С., Информационная РНК и биосинтез белков, М., 1962; Скулачев В. П., Соотношение окисления и фосфорилирования в дыхательной цепи, М., 1962; Ферменты, под ред. А. Е. Браунштейна, М., 1964; Владимиров Г. Е., Пантелеева Н. С., Функциональная биохимия, Л., 1965; И н г р э м В., Биосинтез макромолекул, пер. с англ., М., 1966; Рэкер Э., Биоэнергетические механизмы, пер. с англ., М., 1967; Спирин А. С., Гаврилова Л. П., Рибосома, М., 1968. С. Е. Северин. "БИОХИМИЯ", журнал, издаваемый АН СССР в Москве. Осн. в 1936 А. Н. Бахом. Выходит 1 раз в 2 мес. В журнале публикуются эксперимент, работы по общей биохимии, энзимологии (учение о ферментах), обмену веществ в растениях, животных и микроорганизмах, по биохимии витаминов, гормонов и др. природных физиологически активных соединений, а также по биохимич. основам технологии растительного и животного сырья. Тираж ок. 4 тыс. экз. (1970). О других биохимических журналах см. Биохимия. БИОХОР (от био... и греч. chora -пространство), подразделение биосферы, представляющее собой группу сходных биотопов. Б. объединяются в биоциклы. суша, море и внутр. водоёмы. БИОЦЕНОЗ (от био... и греч. koinos -общий), совокупность растений, животных, микроорганизмов, населяющих участок суши или водоёма и характеризующихся определёнными отношениями как между собой, так и с абиотическими факторами среды (см. Биотоп). Термин "Б." был предложен нем. биологом К. Мёбиусом (1877). Б.- комплекс организмов биогеоценоза, формирующийся в результате борьбы за существование, естественного отбора и других факторов эволюции. По участию в биогенном круговороте веществ в Б. различают три группы организмов. 1) Продуценты (производители) - автотрофные организмы, создающие органич. вещества из неорганических; осн. продуценты во всех Б.- зелёные растения (см. Фотосинтез). Деятельность продуцентов определяет исходное накопление органич. веществ в Б. (см. Биомасса, Биологическая продуктивность). 2) Консументы (потребители)-гетеротрофные организмы, питающиеся за счёт автотрофных. Консументы 1-го порядка - растительноядные животные, а также паразитич. бактерии, грибы и др. бесхлорофильные растения, развивающиеся за счёт живых растений. Консументы 2-го порядка - хищники и паразиты растительноядных организмов. Бывают консументы 3-го и 4-го порядков (сверхпаразиты, суперпаразиты и т. п.), но всего в цепях питания не более 5 звеньев. На каждом последующем трофич. уровне кол-во биомассы резко снижается. Деятельность консументов способствует превращениям и перемещениям органич. веществ в Б., частичной их минерализации, а также рассеянию энергии, накопленной продуцентами. 3) Редуценты (восстановители) - животные, питающиеся разлагающимися остатками организмов (сапрофаги), и особенно непаразитирующие гетеротрофные микроорганизмы - способствуют минерализации органич. веществ, их переходу в усвояемое продуцентами состояние. Взаимосвязи организмов в Б. многообразны. Кроме трофич. связей, определяющих цепи питания (иногда очень своеобразные - см. Паразитизм, Симбиоз), существуют связи, основанные на том, что одни организмы становятся субстратом для других (топические связи), создают необходимый микроклимат и т. п. Часто можно проследить в Б. группы видов, связанные с определённым видом и целиком зависящие от последнего (кон-сорции). Для Б. характерно разделение на более мелкие подчинённые единицы - мероценозы, т. е. закономерно слагающиеся комплексы, зависящие от Б. в целом (напр., комплекс обитателей гниющих дубовых пней в дубраве). Если энергетич. источником Б. служат не автотрофы, а животные (напр., летучие мыши в Б. пещер), то такие Б. зависят от притока энергии извне и являются неполноценными, представляя в сущности мероценозы. В Б. можно выделить и др. подчинённые группировки организмов, напр, синузии. Для Б. также характерно разделение на группировки организмов по вертикали (ярусы Б.). В годовом цикле в Б. изменяются численность, стадии развития и активность отдельных видов, создаются закономерные сезонные аспекты Б. Б.- диалектически развивающееся единство, меняющееся в результате деятельности входящих в него компонентов, вследствие чего происходят закономерные изменение и смена Б. (сукцессии), к-рые могут приводить к восстановлению резко нарушенных Б. (напр., леса после пожара и т. п.). Различают насыщенные и ненасыщенные Б. В насыщенном Б. все экологические ниши (см. Ниша экологическая) заняты и вселение нового вида невозможно без уничтожения или последующего вытеснения к.-л. компонента Б. Ненасыщенные Б. характеризуются возможностью вселения в них новых видов без уничтожения др. компонентов. Можно различать первичные Б., сложившиеся без воздействия человека (целинная степь, девственный лес), и вторичные, изменённые деятельностью человека (леса, выросшие на месте сведённых, население водохранилищ). Особую категорию представляют агробиоценозы, где комплексы осн. компонентов Б. сознательно регулируются человеком. Между первичными Б. и агробиоценозами имеется вся гамма переходов. Изучение Б. важно для рационального освоения земель и водных пространств, т. к. только правильное понимание регулятивных процессов в Б. позволяет человеку изымать часть продукции Б. без его нарушения и уничтожения. Лит.: Кашкаров Д. Н., Основы экологии животных, 2 изд., Л., 1945; Б е к л е м и ш е в В. Н., О классификации биоценологических (симфизиологических) связей, "Бюлл. Московского общества испытателей природы", 1951, т. 56, в. 5;Гиляров М. С., Вид, популяция и биоценоз, "Зоологический журнал", 1954, т. 33, в. 4; А р н о л ь д и К. В. и Л. В., О биоценозе, там же, 1963, т. 42, в. 2; Наумов Н. П., Экология животных, 2 изд., М., 1963; Основы лесной биогеоценологии, под ред. В. Н. Сукачева и Н. В. Дылиса, М., 1964; М а к ф е д ь е н Э., Экология животных, пер. с англ., М., 1965; Одум Е., Экология, М., 1968; Д ю в и н ь о П. и Танг М., Биосфера и место в ней человека, пер. с франц., М., 1968; Tischler W., Synokologie der Landtiere, Stuttg., 1955; Balogh J., Lebensgemeinschaften der Landtiere, Bdpst - В., 1958; Kormondy E. J., Readings in ecology, L., 1965. М. С. Гиляров. БИОЦЕНОЛОГИЯ (от биоценоз я ...логия), наука, изучающая растительные и животные сообщества в их совокупности, т. е. биоценозы, их строение, развитие, распределение в пространстве и во времени, происхождение. Изучение сообществ организмов в их взаимодействии с неживой природой - предмет биогеоценологии. БИОЦЕНОМЕТР (от биоценоз и ...метр), прибор для количеств, учёта наземных насекомых и др. беспозвоночных, применяемый при экологич. исследованиях. Б. представляет собой цилиндр или куб без дна, стенки и верх к-рого затянуты мелкой сеткой или марлей. Применение Б.: пробная площадка размером 0,25-1 м2 быстро (чтобы не ускользнули животные) накрывается Б. Пойманных животных извлекают из Б., умерщвляют, подсчитывают по группам и видам, взвешивают и определяют относит, численность их и массу на единицу площади данного биотопа. БИОЦИКЛЫ, или жизненные области, три самых крупных подразделения биосферы: суша, море и внутр. водоёмы. Каждый Б. подразделяется на биохоры, включающие значит, число биотопов. Напр., биотопы песчаных, глинистых и каменистых пустынь объединяются в биохор пустынь, к-рый вместе с биохорами лесов, степей и др. составляет Б. суши. Термин биологические циклы употребляется в экологии в ином значении. Лит.: Гептнер В. Г., Общая зоогеография, М. -Л., 1936; Наумов Н. П., Экология животных, М., 1955; Вернадский В. И., Химическое строение биосферы Земли и её окружения, М., 1965: Ecological animal geography, N. Y., 1951. И. А. Шилов. БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ, электрические потенциалы, возникающие в тканях и отдельных клетках человека, животных и растений, важнейшие компоненты процессов возбуждения и торможения. Исследование Б. п. имеет большое значение для понимания физико-химич. и физиологич. процессов в живых системах и применяется в клинике с диагностич. целью (электрокардиография, электроэнцефалография, электромиография и др.). Первые данные о существовании Б. п. ("животного электричества") были получены в 3-й четв. 18 в. при изучении природы "удара", наносимого нек-рыми рыбами с электрическими органами при защите или нападении. К этому же времени относится начало исследований итал. физиолога и врача Л. Гальвани, заложивших основу учения о Б. п. Многолетний науч. спор (1791-97) между Л. Гальвани и физиком А. Вольта о природе "животного электричества" завершился двумя крупными открытиями: были получены факты о существовании биоэлектрических явлений в живых тканях и открыт новый принцип получения электрич. тока с помощью разнородных металлов - создан гальванич. элемент (вольтов столб). Правильная оценка наблюдений Гальвани стала возможной лишь после применения достаточно чувствит. электроизмерит. приборов - гальванометров. Первые такие исследования были проведены итал. физиком К. Маттеуччи (1837). Систематич. изучение Б. п. было начато нем. физиологом Э. Дюбуа-Реймоном (1848), к-рый доказал существование Б. п. в нервах и мышцах в покое и при возбуждении. Но ему не удалось (в силу большой инерционности гальванометра) зарегистрировать быстрые, длящиеся тысячные доли сек колебания Б. п. при проведении импульсов вдоль нервов и мышц. В 1886 нем. физиолог Ю. Бернштейн проанализировал форму потенциала действия; франц. учёный Э. Ж. Марей (1875) применил для записи колебаний потенциалов бьющегося сердца капиллярный электрометр; рус. физиолог Н. Е. Введенский использовал (1883) для прослушивания ритмич. разрядов импульсов в нерве и мышце телефон, а гол л. физиолог В. Эйнтховен (1903) ввёл в эксперимент и клинич. практику струнный гальванометр - высокочувствит. и малоинерционный прибор для регистрации электрич. токов в тканях. Значит, вклад в изучение Б. п. внесли рус. физиологи: В. В. Правдич-Немин-ский (1913-21) впервые зарегистрировал электроэнцефалограмму, А. Ф. Самойлов (1929) исследовал природу нервно-мышечной передачи возбуждения, а Д. С. Воронцов (1932) открыл следовые колебания Б. п., сопровождающие потенциал действия в нервных волокнах. Дальнейший прогресс в изучении Б. п. был тесно связан с успехами электроники, позволившими применить в физиологич. эксперименте электронные усилители и осциллографы (работы амер. физиологов Г. Бишопа, Дж. Эрлангера и Г. Гассера в 30-40-х гг. 20в.). Изучение Б.п. в отдельных клетках и волокнах стало возможным с разработкой микроэлектродной техники. Важное значение для выяснения механизмов генерации Б. п. имело использование гигантских нервных волокон головоногих моллюсков, гл. обр. кальмара. Диаметр этих волокон в 50 -100 раз больше, чем у позвоночных животных, он достигает 0,5-1 мм, что позволяет вводить внутрь волокна микроэлектроды, инъецировать в протоплазму различные вещества и т. п. Изучение ионной проницаемости мембраны гигантских нервных волокон позволило англ, физиологам А. Ходжкину, А. Хаксли и Б. Катцу (1947-52) сформулировать современную мембранную теорию возбуждения. Различают следующие осн. виды Б. п. нервных и мышечных клеток: потенциал покоя, потенциал действия, возбуждающие и тормозные постсинаптич. потенциалы, генераторные потенциалы. Потенциал покоя (ПП, мембранный потенциал покоя). У живых клеток в покое между внутр. содержимым клетки и наружным раствором существует разность потенциалов (ПП) порядка 60-90 же, к-рая локализована на поверхностной мембране.
Рис. 2. Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов: а - гигантского аксона кальмара; 6 - скелетного мышечного волокна; в - волокна мышцы сердца собаки; 1 - восходящая фаза ПД; 2 - нисходящая фаза; 3 - следовая гиперполяризация (а) и следовая деполяризация (б). Внутр. сторона мембраны заряжена электроотрицательно по отношению к наружной (рис. 1). ПП обусловлен избират. проницаемостью покоящейся мембраны для ионов К+ (Ю. Бернштейн, 1902, 1912; А. Ходжкин и Б. Катц, 1947). Концентрация К+ в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутр. сторона мембраны, практически не проницаемой для крупных органич. анионов, приобретает отрицат. потенциал. Поскольку проницаемость мембраны в покое для Na+ примерно в 100 раз ниже, чем для К+, диффузия натрия из внеклеточной жидкости (где он является осн. катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К+. В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы С1-, диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между повреждённым и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na+ и К+ падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na+ значительно превышает скорость увеличения проницаемости мембраны для К+.
Рис. 1. Схема измерений мембранного потенциала покоя с помощью внутриклеточного стеклянного микроэлектрода (М). Второй электрод (И) помещён в омывающую клетку жидкость. Потенциал действия (ПД). Все раздражители,
действующие на клетку, вызывают в первую очередь снижение ПП; когда оно
достигает критич. значения (порога), возникает активный распространяющийся
ответ - ПД (рис. 2). Во время восходящей фазы ПД кратковременно извращается
потенциал на мембране: её внутр. сторона, заряжённая в покое электроотрицательно,
приобретает в это время положит, потенциал. Достигнув вершины, ПД начинает
падать (нисходящая фаза ПД), и потенциал на мембране возвращается к уровню,
близкому к исходному, - ПП. Полное восстановление ПП происходит только
после окончания следовых колебаний потенциала - следовой деполяризации
или гиперполяризации, длительность к-рых обычно значительно превосходит
продолжительность пика ПД. Согласно мембранной теории, деполяризация мембраны,
вызванная действием раздражителя, приводит к усилению потока Na+
внутрь клетки, что уменьшает отрицат. потенциал внутр. стороны мембраны
- усиливает её деполяризацию, Это, в свою очередь, вызывает дальнейшее
повышение проницаемости для Na+ и новое усиление деполяризации
и т. д.
Рис. 3. Изменения натриевой и калиевой проводимости мембраны нервного волокна во время генерации потенциала действия (I). Изменения проводимости пропорциональны изменениям проницаемости для Na+ (II) и К+ (III). В результате такого взрывного кругового процесса, т. н. регенеративной деполяризации, происходит извращение мембранного потенциала, характерное для ПД. Повышение проницаемости для Na+ очень кратковременно и сменяется её падением (рис. 3), а следовательно, уменьшением потока Na+ внутрь клетки. Проницаемость для К+, в отличие от проницаемости для Na+, продолжает увеличиваться, что приводит к усилению потока К+ из клетки. В результате этих изменений ПД начинает падать, что ведёт к восстановлению ПП. Таков механизм генерации ПД в большинстве возбудимых тканей. Существуют, однако, клетки (мышечные волокна ракообразных, нервные клетки у ряда брюхоногих моллюсков, нек-рые растит, клетки), у к-рых восходящая фаза ПД обусловлена повышением проницаемости мембраны не для ионов Na+, а для ионов Са2+. Своеобразен также механизм генерации ПД в мышечных волокнах сердца, для к-рых характерно длительное плато на нисходящей фазе ПД (рис. 2, в). Неравенство концентраций ионов К+ и Na4+ (или Са2+ ) внутри и снаружи клетки (волокна) поддерживается специальным механизмом (т. н. "натриевым насосом"), выталкивающим ионы Na+ из клетки и нагнетающим ионы К+ в протоплазму, требующим затраты энергии, которая черпается клеткой в процессах обмена веществ. Амплитуда ПД большинства нервных и мышечных волокон примерно одинакова: 110-120 мв. Длительность ПД варьирует в широких пределах: у теплокровных животных длительность ПД нервных волокон, наиболее быстро проводящих возбуждение, -0,3-0,4 мсек, у волокон же мышц сердца - 50-600 мсек. В растит, клетках пресноводной водоросли хара ПД продолжается ок. 20 сек. Характерной особенностью ПД, отличающей его от др. форм ответа клетки на раздражение, является то, что он подчиняется правилу "всё или ничего", т. е. возникает только при достижении раздражителем нек-рого порогового значения, и дальнейшее увеличение интенсивности раздражителя уже не сказывается ни на амплитуде, ни на продолжительности ПД, Потенциал действия - один из важнейших компонентов процесса возбуждения. В нервных волокнах он обеспечивает проведение возбуждения от чувствит. окончаний (рецепторов) к телу нервной клетки и от неё - к синаптическим окончаниям (см. Синапсы), расположенным на различных нервных, мышечных или железистых клетках. Поступая в эффекторные окончания, ПД вызывает выделение (секрецию) определённой порции специфич. хим. веществ, т. н. медиаторов, оказывающих возбуждающее или тормозящее влияние на соответств. клетки. В мышечных волокнах распространяющийся ПД вызывает цепь физико-химич. реакций, лежащих в основе процесса сокращения мышц. Проведение ПД вдоль нервных и мышечных волокон осуществляется т. н. локальными токами, или токами действия, возникающими между возбуждённым (деполяризованным) и соседними с ним покоящимися участками мембраны (см. Возбуждение). Токи действия регистрируются обычными внеклеточными электродами; при этом кривая имеет двухфазный характер: первая фаза соответствует приходу ПД под ближний электрод, вторая - под дальний электрод (рис. 4).
Рис. 4. Регистрация распространения потенциала действия вдоль нервного волокна. А, Б - внеклеточные электроды; р - раздражающие электроды. Вверху - отклонение луча осциллографа под влиянием волны возбуждения; 1 - волна возбуждения под электродом А; 2 - под электродом Б. Постсиваптические потенциалы (ПСП) возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мв и длительность 10-15 мсек. ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД (рис. 5, а, 6). ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора (рис. 5, в). В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки.
Рис. 5. Постсинаптические потенциалы: а - подпороговые ВПСП, возникающие в нервной клетке в ответ на раздражения соответствующих нервных волокон; б - ВПСП, достигший порогового значения, достаточного для возникновения ПД; в - ТПСП, вызванный раздражением тормозных нервных волокон. Гевераторвые потенциалы возникают в мембране чувствит. нервных окончаний - рецепторов. Они внешне сходны с ВПСП - их амплитуда порядка нескольких мв и зависит от силы приложенного к рецептору раздражения (рис. 6). Когда генераторный потенциал достигает порогового (критического) значения, в соседнем участке мембраны нервного волокна возникает распространяющийся ПД. Ионный механизм генераторных потенциалов ещё недостаточно изучен. Наряду с перечисленными относительно быстро развивающимися Б. п., в нервных клетках, волокнах гладких мышц и нек-рых растит, клетках регистрируются также очень медленные колебания мембранного потенциала неизвестной природы, причём на гребне волны деполяризации мембраны часто возникают разряды импульсов.
Рис. 6. Генераторные потенциалы; увеличение амплитуды при усилении раздражения рецептора (а - е). При достижении порогового значения (в) генераторный потенциал вызвал в чувствительном нервном волокне потенциал действия. Все Б. п. могут быть зарегистрированы В точно измерены только с помощью внутриклеточных микроэлектродов, позволяющих отводить разности потенциалов между внутренней и наружной сторонами мембраны клетки. При отведении колебаний Б. п. от целых нервов, мышц или мозга с помощью поверхностных электродов регистрируется лишь суммарно потенциал множества синхронно или, чаще, асинхронно работающих клеток. Так, электромиограмма представляет собой результат сложения (интерференции) ПД множества скелетных мышечных волокон; электрокардиограмма - результирующая колебаний электрич. потенциалов мышечных волокон различных отделов сердца; электроэнцефалограмма -результат суммации гл. обр. ВПСП и ТПСП множества клеток различных слоев коры больших полушарий. Регистрация таких интерференционных электрограмм, хотя и не позволяет анализировать колебания Б. п. отдельных клеток, имеет важное значение для суждения о состоянии исследуемого органа в целом. В клинич. практике электромиограмму, электрокардиограмму и электроэнцефалограмму регистрируют с помощью электродов, расположенных на коже соответств. частей тела. Оценка данных, полученных этими методами, основана на сопоставлении изменений характера соответств. кривой с результатами клинич., физиологич. и патологоанатомич. исследований. Лит.: физиология человека, М., 1966; Гальвэни Л. и Вольта А., Избр. работы о животном электричестве, М. - Л., 1937; Ходжкин А., Нервный импульс, пер. с англ., М., 1965; Э к к л с Д ж.. Физиология нервных клеток, пер. с англ., М., 1959; его же, физиология синапсов, М., 1966; К а т ц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; ХодоровБ. И., Проблема возбудимости, Л., 1969. Б.И. Ходоров. БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ, см. Биоэлектрические потенциалы. БИОЭНЕРГЕТИКА, биологическая энергетика, изучает механизмы преобразования энергии в процессах жизнедеятельности организмов. Иначе говоря, Б. рассматривает явления жизнедеятельности в их энергетич. аспекте. Методы и подходы к изучаемым явлениям, применяемые в Б.,- физико-химические, объекты и задачи - биологические. Т. о., Б. стоит на стыке этих наук и является частью молекулярной биологии, биофизики и биохимии. Началом Б. можно считать работы нем. врача Ю. Р. Майера, открывшего закон сохранения и превращения энергии (1841) на основе исследования энергетич. процессов в организме человека. Суммарное изучение процессов, являющихся источниками энергии для живых организмов (см. Дыхание, Брожение), и энергетич. баланса организма, его изменений при различных условиях (покой, труд разной интенсивности, окружающая темп-ра) долгое время являлось основным содержанием Б. (см. Основной обмен, Теплоотдача, Теплопродукция). В середине 20 в., в связи с общим направлением развития биологич. наук, центральное место в Б. заняли исследования механизма преобразования энергии в живых организмах. Все исследования в области Б. основываются на единственно науч. точке зрения, согласно к-рой к явлениям жизни полностью применимы законы физики и химии, а к превращениям энергии в организме - осн. начала термодинамики. Однако сложность и специфичность биологич. структур и реализующихся в них процессов обусловливают ряд глубоких различий между Б. и энергетикой неор-ганич. мира, в частности технич. энергетикой. Первая фундаментальная особенность Б. заключается в том, что организмы - открытые системы, функционирующие лишь в условиях постоянного обмена веществом и энергией с окружающей средой. Термодинамика таких систем существенно отличается от классической. Основополагающее для классич. термодинамики понятие о равновесных состояниях заменяется представлением о стационарных состояниях; второе начало термодинамики (принцип возрастания энтропии) получает иную формулировку в виде Пригожина теоремы. Вторая важнейшая особенность Б. связана с тем, что процессы в клетках протекают в условиях отсутствия перепадов темп-ры, давления и объёма; в силу этого переход теплоты в работу в организме невозможен и тепловыделение представляет невозвратимую потерю энергии. Поэтому в ходе эволюции организмы выработали ряд специфич. механизмов прямого преобразования одной формы свободной энергии в другую, минуя её переход в тепло. В организме лишь небольшая часть освобождающейся энергии превращается в тепло и теряется. Большая её часть преобразуется в форму свободной хим. энергии особых соединений, в к-рых она чрезвычайно мобильна, т. е. может и при постоянной темп-ре превращаться в иные формы, в частности совершать работу или использоваться для биосинтеза с весьма высоким кпд, достигающим, напр, при работе мышцы, 30%. Одним из осн. результатов развития Б. в последние десятилетия является установление единообразия энергетических процессов во всём живом мире - от микроорганизмов до человека. Едиными для всего растит, и животного мира оказались и те вещества, в к-рых энергия аккумулируется в подвижной, биологически усвояемой форме, и процессы, с помощью к-рых такое аккумулирование осуществляется. Такое же единообразие установлено и в процессах использования аккумулированной в этих веществах энергии. Например, структура сократит, белков и механизм механо-химич. эффекта (т. е. превращения химич. энергии в работу) в основном одни и те же при движении жгутиков у простейших, опускании листиков мимозы или при сложнейших движениях птиц, млекопитающих и человека. Подобное единообразие характерно не только для явлений, изучаемых Б., но и для др. присущих всему живому функций: хранения и передачи наследств, информации, осн. путей биосинтеза, механизма ферментативных реакций. Веществами, через к-рые реализуется энергетика организмов, являются макроэргические соединения, характеризующиеся наличием фосфатных групп. Роль этих соединений в процессах превращения энергии в организме впервые установил, изучая мышечное сокращение, сов. биохимик В. А. Энгелъгардт. В дальнейшем работами мн. исследователей было показано, что эти соединения участвуют в аккумуляции и трансформации энергии при всех жизненных процессах. Энергия, освобождающаяся при отщеплении фосфатных групп, может использоваться для синтеза биологически важных веществ с повышенным запасом свободной энергии и для процессов жизнедеятельности, связанных с превращением свободной химич. энергии в работу (механическую, активного переноса веществ, электрическую и т. д.). Важнейшим из этих соединений веществом, играющим для всего живого мира роль почти единственного трансформатора и передатчика энергии, является аденозинтрифосфорная к-та - АТФ (см. Аденозинфосфорные кислоты), расщепляющаяся до аденозиндифосфорной к-ты (АДФ)или аденозинмонофосфорной к-ты (АМФ). Гидролиз АТФ, т. е. отщепление от неё конечной фосфатной группы, протекает по уравнению: АТФ + Н2О -> АДФ + фосфат и сопровождается уменьшением свободной энергии на значение дельта F. Если эта реакция протекает при концентрации всех реагентов и продуктов в 1,0 моль при 25°С и рН 7,0, то свободная энергия АДФ оказывается меньше свободной энергии АТФ на 29,3 кдж. (7000 кал). В клетке это изменение свободной энергии больше: дельта F = 50 кдж/моль (12 000 кал/моль). Значения ДР для реакции АТФ->АДФ выше, чем у большинства реакций гидролиза. Макроэргическими называют и сами связи третьей (конечной) и второй фосфатных групп в молекуле АТФ и аналогичные связи в др. макроэргических соединениях. Эти связи обозначают знаком ~ (тильда); напр., формулу АТФ можно записать так: аденин - рибоза -фосфат ~ фосфат ~ фосфат. Говоря об энергии макроэргических связей, в Б. имеют в виду не действительную энергию ковалентной связи между атомами фосфора и кислорода (или азота), как это принято в физич. химии, а лишь разность между значениями свободной энергии (дельта F) исходных реагентов и продуктов реакций гидролиза АТФ или др. аналогичных реакций. "Энергия связи" в этом смысле, строго говоря, не локализована в данной связи, а характеризует реакцию в целом. Энергия макроэргических связей АТФ является универсальной формой запасания свободной энергии для всего живого мира: все преобразования энергии в процессах жизнедеятельности осуществляются через аккумуляцию энергии в этих связях и её использование при их разрыве. Значение дельта F для этих реакций представляет собой как бы "биологический квант" энергии, т. к. все преобразования энергии в организмах происходят порциями, примерно равными дельта F. При ферментативном гидролизе АТФ в клетке отщепляющаяся фосфатная группа всегда переносится на субстрат, запас энергии в к-ром оказывается в результате больше, чем в исходном соединении. Обмен веществ (метаболизм) в клетке состоит из непрерывно совершающихся распада сложных веществ до более простых (катаболические процессы) и синтеза более сложных веществ (анаболические процессы). Катаболические процессы являются экзергоническими, т. е. идут с уменьшением свободной энергии (дельта F<0); анаболические процессы -эндергонические, они протекают с увеличением свободной энергии (дельта F>0). Согласно общим законам термодинамики, экзергонич. процессы могут протекать спонтанно, самопроизвольно, процессы же эндергонические требуют притока свободной энергии извне. В клетке это осуществляется благодаря сопряжению обоих процессов: одни используют энергию, освобождаемую при протекании других. Это сопрялсение, лежащее в основе всего метаболизма и жизнедеятельности клетки, совершается при посредстве системы АТФ - АДФ, создающей промежуточные, обогащённые энергией соединения. Напр., синтез сахарозы из глюкозы и фруктозы происходит за счёт энергии, освобождающейся при реакции гидролиза АТФ, путём образования промежуточного активированного соединения - глю-козо-1-фосфата: 1) АТФ + глюкоза -> -> АДФ + глюкозо-1-фосфат; 2) глюкозо-1-фосфат + фруктоза -> сахароза + фосфат. Суммарная реакция: АТФ + глюкоза + фруктоза -> АДФ + сахароза + фосфат. Энергетич. баланс процесса: АТФ->АДФ + фосфат - 29,3 кдж/моль (-7000 кал/моль) (уменьшение свободной энергии); глюкоза + фруктоза -> сахароза + 23 кдж/моль ( + 5500 кал/моль) (увеличение свободной энергии). Потеря энергии на тепло 6,3 кдж/моль (1500 кал/моль), т. е. кпд процесса 79%. По такому же типу осуществляется сопряжение реакций и при синтезе др. сложных соединений (липидов, полисахаридов, белков и нуклеиновых к-т). В этих процессах, кроме АТФ, принимают участие и нек-рые аналогичные соединения, в к-рые, вместо аденина, входят др. азотистые основания (гуанин-, цитозин-, уридин-, тимидинтрифосфаты или креатинфосфаты). При синтезе белков и нуклеиновых к-т от АТФ отщепляется не одна концевая фосфатная группа, а две последние (пирофосфат). Т. о., все процессы накопления (аккумулирования) энергии в организмах должны сводиться к процессам образования АТФ, т. е. фосфорилирования (включения фосфатных групп в АДФ или АМФ). Энергетика процессов метаболизма, в к-рых энергия сохраняет форму химической, в основных чертах ясна, но этого нельзя сказать о процессах, в к-рых энергия переходит из химич. формы в механич. работу или какой-нибудь иной вид энергии (напр., электрический). Так, известно, напр., что работа, совершаемая сокращающейся мышцей, производится за счёт энергии, освобождающейся при гидролизе АТФ, но механизм этого преобразования энергии ещё не ясен. Выяснение интимных механизмов механохимич. эффекта и др. превращений химич. энергии - важная и актуальная задача Б., успешное решение к-рой может открыть путь к прямому преобразованию химич. энергии в механическую и электрическую без промежуточного "разорительного" превращения её в тепло. Осн. и практически единственным источником энергии для жизни на Земле является энергия излучения Солнца, часть к-рой поглощается пигментами растений и нек-рых бактерий и в процессе фотосинтеза аккумулируется автотрофными организмами в форме химич. энергии: частью в виде АТФ (процессы фотосинтетич. фосфорилирования), частью в виде энергии нек-рых специфич. соединений (восстановленных никотин-амид-адениндинуклеотидов), являющихся важнейшими промежуточными аккумуляторами энергии. Весь дальнейший процесс синтеза углеводов, а затем и липидов, белков и др. компонентов клетки осуществляется в цикле темновых ферментативных реакций за счёт энергии указанных выше соединений. При реакции синтеза углеводов [суммарно: 6СО2+6Н2О -> C6Hi2O6 + 6O2] увеличение свободной энергии дельта F=2,87 Мдж/моль (686 000 кал/моль), а теплосодержание продуктов (молярная энтальпия) изменяется на величину дельта Н=2,82 Мдж/молъ (673 000 кал/моль). Т. о., углеводы, липиды, белки и др. пищевые продукты представляют собой форму долговременного хранения поглощённой растением энергии излучения. В гетеротрофных организмах АТФ образуется в процессе дыхания на промежуточных стадиях окисления пищевых веществ до СО2 и воды. В этом процессе ок. 40-50% свободной энергии переходит в энергию макроэргич. связей АТФ, а остальная теряется в виде тепла. Общее кол-во энергии, запасаемой растениями в год (при упрощённом предположении, что весь углерод фиксируется в виде глюкозы), равно примерно 1018-1021дж, что составляет лишь 0,001 от общего потока падающей на Землю солнечной энергии (1024 дж/год). Нек-рое кол-во энергии накапливается и в процессах хемосинтеза за счёт окисления восстановленных неорганич. соединений, но вклад этих процессов в энергетику биосферы невелик. Сказанное выше характеризует только суммарный баланс энергии в процессах её аккумуляции и использования. Изучение первичных механизмов миграции энергии на клеточном и молекулярном уровнях показало, что решающую роль в них играет транспорт электронов по цепи передатчиков. В отдельных звеньях этой цепи окислительно-восстановительных реакций происходит освобождение небольших порций свободной энергии, примерно соответствующих значениям дельта F для макроэргич. связей АТФ. Дальнейшее изучение проблем Б., в частности механизмов преобразования химич. энергии в работу, требует перехода к рассмотрению этих процессов на субмолекулярном уровне, где вступают в силу законы квантовой физики и химии. Лит.: Виноградов М. И., Очерки по энергетике мышечной деятельности человека, Л., 1941; Сент-Дьердьи А., Биоэнергетика, пер. с англ., М., 1960; его же, Введение в субмолекулярную биологию, пер. с англ., М., 1964; П а с ы н с к и й А. Г., Биофизическая химия, М., 1963; Горизонты биохимии. Сб. ст., под ред. Л. А. Тумермана, пер. с англ., М., 1964; Пюльман Б., Пюльман А., Квантовая биохимия, пер. с англ., М., 1965; Ленинджер Л., Митохондрия, пер. с англ., М., 1966; Л е м а н Г., Практическая физиология труда, пер. с нем., М., 1967; Рэкер Э., Биоэнергетические механизмы, пер. с англ., М., 1967; Lеhninger A. L., Bioenergetics, N. Y., 1965; Current topics in bioenergetics, ed. D. R. Sa-nadi, v. 1 - 2, N. Y., 1966-67. Л. А. Тумерман.
|